In the first section of this three part series, we examined the background and defining characteristics of the Late Quaternary Extinctions and explored the nuances of climate change and ecology. In doing so, we showed that the support for climate as the central driver of the disappearance of so many species over the past few tens of thousands of years is very weak, making climate-dominant positions untenable. However, as mentioned previously, there is a popular and seemingly intuitive notion that climate change and human agency played roughly equivalent roles.
While this stance seems reasonable at first glance, we need to take a step back and reevaluate whether the evidence actually supports it or if this is simply an argument to moderation fallacy. It is undeniable that the climate was changing to one extent or another during many extinction waves, sometimes dramatically so. Yet this can be something of a red herring because the climate is always changing, and as humans arrived on various continents at different times, their arrival often coincided with substantial shifts in climate by chance alone. As the previous section demonstrated, we cannot automatically assume that climatic change, by virtue of occurring simultaneously with extinctions on a given continent, necessarily played a meaningful part in driving them.
So how do we decide how important climate actually was in relation to humans? It is tempting to believe that disentangling the impacts of climate change and humans on megafauna is inherently difficult or even impossible given that both were frequently co-occurring. Fortunately, it is not a futile endeavor—analyzing extinction patterns through biogeography offers an effective way to do just that.
Biogeography
To give us a clue as to whether climate was roughly as important as humans or much less so, we can look at extinction rate by continent and compare it to the magnitude of climate change. A basic expectation in a scenario where climate featured roughly as prominently as humans in extinction would be that a strong link still emerges between extinction severity and climate change magnitude, with continents that experienced the greatest degree of climate change such as North America and Europe also having much greater extinction rates than continents with milder climate change, like South America or Australia.
Yet, when we actually compare the various continents by these metrics, something very odd happens: studies find the link between climate change magnitude and extinction severity to be either weak or absent1 2 3. Instead, a far stronger inverse correlation emerges between the severity of extinction and the duration of hominin presence in a region, while the timing of extinction waves is closely tied to the arrival of anatomically modern humans.
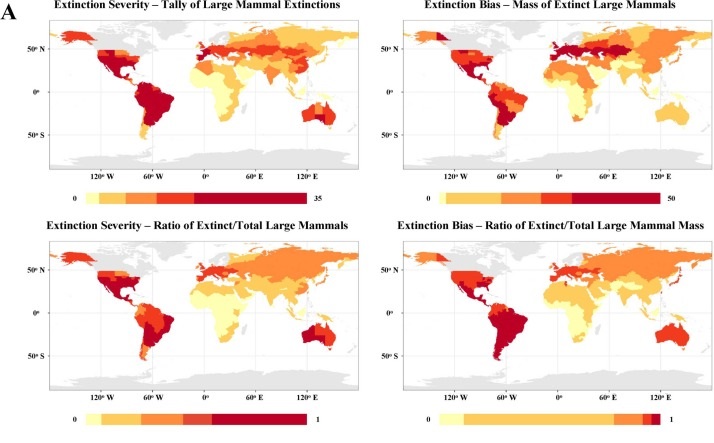
A) Extinction severity along various metrics
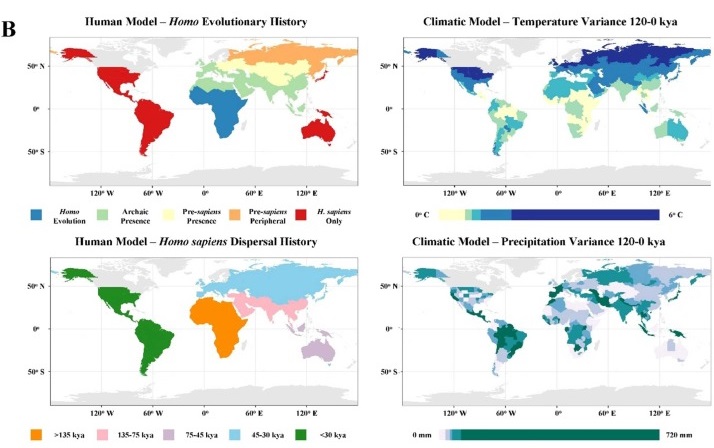
B) Human history and climate change metrics
This has major implications for the extinction debate, yet there is an often overlooked element of this sort of global analysis that can give us substantial insight: what occurred specifically on the three continents without any prior hominin presence—North America, South America, and Australia (then part of Sahul).
The New World and Sahul
These three continents provide an ideal natural experiment because they each had megafauna that were entirely unfamiliar with humans but experienced climate change very differently owing to their distinct geographical positions. The proposed climatic mechanisms for North America, South America, and Australia involve, respectively, deglaciation-linked climate shocks4, loss of open-habitat (in conjunction with human pressure) 5, and protracted aridification6. The flaws in each of these theories are legion and I encourage reading my previous posts dedicated to dismantling them for those who would like more information: (1)(2).

However, for the purpose of this article, I would like to focus on what comparing the extinction rates between the three can tell us: the proportion of megafaunal losses goes from highest to lowest in Australia (90%), South America (83%), and finally North America (72%) 7. Why is that significant? Because this pattern is exactly the opposite of what one would expect if climate played a substantial role in extinctions in them, and precisely what we would expect to see if it was humans! Let me explain.
For a bit of background, extinctions in North America (south of the erstwhile ice sheet) and South America were more or less contemporaneous8. It has been claimed that the extinction process took much longer in South America than in North America9 10 but this notion is based mainly on remains of allegedly late surviving megafauna from the Pampas, the dates for which have already been shown to be erroneously young11. The extinction window for megafauna in South America was considerably narrowed down in a landmark study from 202112 and even further in a newer study from 202513. In other words, we can reasonably set the megafaunal decline across the Americas to the Pleistocene-Holocene transition.
The severity of climate change experienced during the respective extinction windows of the aforementioned three continents goes in the order of North America>South America>Australia. North and South America both were in the midst of a glacial-interglacial transition when extinctions occurred, but North America had more extreme climate change. Rapid temperature fluctuations over the course of decades or centuries compounded pronounced continentality during this transition in much of North America14 unlike in South America. Meanwhile, extinctions in Australia occurred tens of thousands of years earlier during a time of mild climate15 16.
Therefore, the correlation with climate change is actually negative here, which is seriously problematic for those who would argue, for instance, that the high extinction rates in North and South America were due to human arrival coinciding with a glacial termination17 18. However, this pattern can be explained perfectly well with primarily human agency. Recall how we mentioned earlier that the extinctions were least severe in Africa and most severe on islands; Africa is the cradle of our species and hence where wildlife was best acquainted with man while islands had naive faunas lacking formidable predators. While North America, South America, and Australasia are not known to have had any hominin habitation prior to Homo sapiens arrival, there is still a key difference in terms of how “island-like” and “non-African” they are/were in terms of their megafaunal assemblage, and hence their vulnerability to human pressure.
North America was the least island-like, having been connected periodically to the Eurasian (and by extension African) landmass for tens of millions of years. There was extensive biotic interchange as animals crossed Beringia, but the traffic tended to go more from west to east than vice versa. By the Late Pleistocene, North America had a particularly large guild of ferocious predators, and this meant that prey species likely developed relatively stronger defenses (greater speed, intelligence, and/or reproductive rates) than their South American and especially Australian counterparts19.
South America, on the other hand, was far more isolated as it was an island for tens of millions of years. That isolation was broken by the formation of the Isthmus of Panama 2.7 million years ago, and this allowed North American forms to establish themselves on the continent, including lethal predators. This linkage could have partially given South American animals an advantage relative to Australian ones when it came to surviving the eventual human onslaught, but they almost certainly still lagged behind North American counterparts in that regard given the previous lengthy period of isolation. Another aspect is that herbivores—disproportionately represented by the Xenarthran superorder—in South America tended to be larger than North American ones, which could increase extinction risk as larger animals were more vulnerable to humans.

Australia was (and still is) by far the most insular: it has been disconnected from other continents for tens of millions of years, and marsupial forms dominate. There were important terrestrial reptilian predators like Quinkana and Megalania, which may seem terrifying to casual observers, but by the Quaternary, these forms could only have persisted in marsupial-dominated Australia as they could not have competed with placental carnivores like tigers and lions in places like Africa and Asia19. Predators who were less formidable meant less developed anti-predatory defenses among prey, which meant easy pickings for a superpredator like Homo sapiens.

While it is impossible to plainly state that climate change played no role whatsoever in extinction dynamics on these three continents, the patterns seen here certainly cannot be reconciled with it having a substantial part in them. But this begs the question of why this might be the case. After all, even if humans were a highly destructive force, why would climate only have a minor imprint on these extinction patterns, even in North America? The answer, I would argue, is fairly simple: the ability to survive climate change is highly dependent on, among other things, the ability of animals to disperse geographically.
I wrote in the first part of my series on American extinctions about how North and South America largely lack major mountainous barriers east of the Western mountain ranges (Rockies, Andes, etc.) which would allow animals to move from wet/forested areas to drier/grassier areas with relative ease. Likewise, Australia has its own version of this where topographic barriers between the more humid coasts and arid interior are not significant, with even the Great Dividing Range not being particularly tall.
Therefore, freedom of movement would have provided animals an effective way to cope with climate change, large or small, on these three continents; this would explain why climate change appears to not be a very significant factor there. The role of geographic barriers is underrated and will be an important point as we discuss extinctions in Europe and Asia in the next section.
Eurasia-Northern vs. Southern
In Eurasia (Europe and Asia), where there was an extended presence of archaic humans such as Neanderthals, Denisovans, Homo heidelbergensis, and Homo erectus prior to anatomically modern humans, the situation is somewhat more complex. Studies that look at the LQE from a global standpoint only identify either an unclear or weak link between extinction and climate change on that continent1 2. However, studies that focus on Eurasia specifically tend to be dominated by climate-based extinction models20. It is therefore important to take a closer look at Eurasia to see if we can reconcile this discrepancy—could it be that the finer points of climate-megafauna interactions there are being missed by global-scale analysis?
The extinctions in Eurasia were staggered, starting after the initial human expansion out of Africa 60 to 50 thousand years ago and extending well into the Holocene21. In many cases, the timing of the extinction of respective taxa, to an extent, made sense. For example, temperate-adapted animals went extinct during the glacial period whereas mammoth steppe animals went extinct much later, in the late glacial or even Holocene22. Furthermore, southern Eurasia has retained more of its overall megafaunal diversity, and especially large herbivores, than the more environmentally turbulent northern Eurasia1 21. While the latter has often been explained through hominin biogeography, there are problems with this interpretation as hominins, in this case Neanderthals and Denisovans, were present in much of mid-latitude northern Eurasia (between roughly 40-50 degrees north) for an extended period of time.
Therefore, we can take a closer look at what might be happening in Eurasia. The landmass features a highly complex geography including west-east mountain ranges, a dizzying arrangement of vegetation zones, and several inland/marginal seas; this is all in stark contrast to the much more simplified geography of the New World and Australia. This creates difficulty for the dispersal of plants and animals. Extinctions or bottlenecks are more likely when those animals forced to move to refugia when the climate changes in an adverse way.
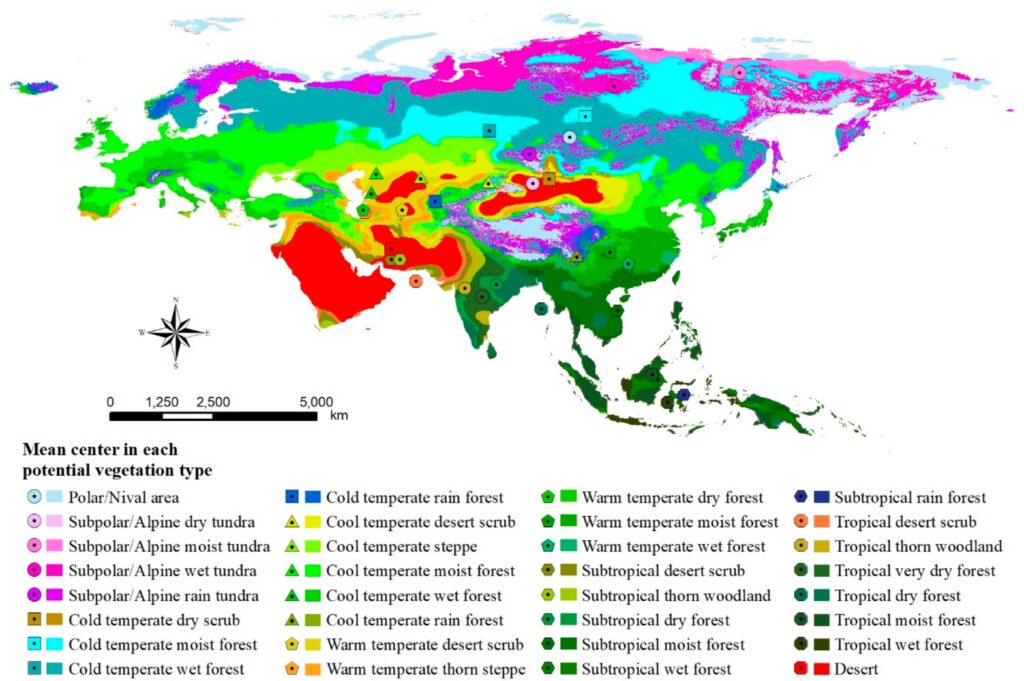
Europe is a particularly challenging milieu for dispersal, with mountains and water bodies impeding passage between northern, central, and southern Europe and also separating the continent from West Asia and North Africa. In contrast to low latitudes where climatic shifts and faunal turnovers were not as severe, mid and upper latitudes experienced great change; Europe oscillated between very cold glacial climates dominated by steppic environments to much more mild interglacial climates dominated by temperate forests and woodlands23. For a temperate-climate animal in central Europe trying to survive glaciation, moving to the Mediterranean peninsulas could be a solution. However, mountains like the Alps and Pyrenees would need to be bypassed.
Forests in general become rare throughout Europe and Siberia during glacial maxima and give way to steppe-tundra24. This would obviously be problematic for animals that depend on said forests. With the onset of interglacials meanwhile, heavy afforestation takes place from the Atlantic all the way to the Pacific coasts of Eurasia, and it is possible that many steppe-tundra animals may get trapped by being too far away to reach the Central Asian steppes or Arctic tundra in time.
Because of these realities, it is tempting to speculate that the relative importance of climatic fluctuation may be greater in Eurasia. Again, extinctions were more severe in northern Eurasia than in southern Eurasia, even in areas where archaic hominins such as Neanderthals and Denisovans coexisted with native megafauna. Extinctions were staggered and often coincident with environmental stress for particular species. This could indicate a possible role for climate. Even so, it would be quite strange to argue that extinctions in Eurasia were entirely or mostly climatic in origin, because the nature of the losses is so striking and unprecedented.
Four genera of proboscideans comprising several species were present on the continent prior to anatomically modern human expansion: Mammuthus (including the woolly mammoth, M. primigenius), Paleoloxodon (the straight-tusked elephants, with multiple species across Eurasia), Stegodon (various species throughout Asia), and the Asian elephant, Elephas maximus. Only the last is still extant. The rhinoceros guild was far more widespread and diverse too, but now restricted solely to tropical southern and southeastern Asia. This, again, is too drastic of a change to be attributable to climate alone.


Additionally, patterns of extinction in Eurasia may also be partly explained by human-population dynamics rather than environmental effects on fauna alone. Harsh glacial conditions likely delayed widespread Homo sapiens settlement in higher latitudes of the mammoth steppe until the onset of warming25. The scarcity of trees and (to a lesser extent) shrubs was a common feature of these northerly regions. This would have limited the availability of wood, which is so essential to humans for firemaking and tool production26.
As a result, the modern human population of the northern half of Siberia and the part of Europe adjacent to the erstwhile ice sheet—intensely cold places where archaic human presence was also absent or very limited—was constrained in distribution and density for an extended period of time. That is, until the glacial–interglacial transition between 15 and 11 thousand years ago allowed woody vegetation, and hence human habitation, to expand in these areas to an extent that was unprecedented in the preceding tens of thousands of years. This may partially explain the lengthy persistence of mammoth steppe fauna long after initial human entry into northern Eurasia and the timing of their ultimate demise.
Moving now to southern Eurasia, there is a possibility that extinction rates here were lower than those in the continent’s northerly regions because tropical diseases, as in Africa, may have limited human population density during the Paleolithic27. This remains speculative, but if true, this would be another indication that the direct effects of climate change on Eurasian megafauna were less significant than they are often assumed to be. Climate may have influenced Eurasian extinction patterns more indirectly through its controls on the human population.
Africa
In Africa, where anatomically modern humans emerged and spent the longest amount of time, losses were only truly significant in North Africa while Sub-Saharan Africa was mainly spared1 21. In Sub-Saharan Africa, Late Quaternary megafauna which went extinct prior to the colonial period are Syncerus antiquus (the giant long-horned buffalo), Megalotragus priscus (giant wildebeest/hartebeest), and Equus capensis (the Cape zebra). The status of the last as its own species has sometimes been contested28, but it was certainly much larger than living zebras. The exact causes of the handful of extinctions in Sub-Saharan Africa are unknown but it is clear that no niches are vacant as close relatives of these species are still present.
North Africa is a complex case. African humid periods occur cyclically and are known to cause a proliferation of savannah in what is now the bone-dry Sahara desert; animals typical of Sub-Saharan Africa such as giraffes, hippos, elephants, antelopes, and others were periodically widespread, as indicated by their depictions in cave art29. As the last humid period ended around the mid-Holocene, North African megafauna contracted their ranges into refugia such as the Nile River or Mediterranean coast, or moved south. Despite the drastic environmental change, many of them only went extinct in the Late Holocene (in some cases only the last few centuries) meaning that humans were certainly involved in their final extinctions. Moreover, most of these losses could be better described as extirpations rather than extinctions as their conspecifics still exist just south of the Sahara.


Now we can put all of this together. Extinctions were most severe in the three continents without prior hominin presence, and given the fact that extinction intensity does not align with the strength of climate change, we are left to conclude that humans played the dominant, overriding force there. Extinctions in Eurasia were less severe but still unprecedented, which means humans had to have had a substantial hand, even if climate contributed to one extent or another. And of course, humans were the sole cause of extinctions on the islands. The math here is not hard: globally, humans were necessarily more important than climate change by far when it comes to the LQE.
Having finally established this (for good), we can move on to discussing why it is that early humans were so destructive to the environments and animal communities they encountered.
Man the Juggernaut
There are many adjectives that can be used to describe the picture of early humans moving continent to continent wiping out masses of animals as they went: morbid, strange, unsettling. But perhaps the best descriptor is shocking. Many people look at how countless species of animals in the modern day continue to cling on, even thrive, despite humans having acquired advanced hunting weapons like guns and then are baffled wondering about how much smaller numbers of Paleolithic humans with far more primitive weapons could have dealt such devastating damage to animal communities of the past. However, we must keep in mind that we are looking at a world with a profoundly shifted baseline in terms of vulnerability to humans; species that still exist now are ones that successfully made it past a highly selective filter.
It is notable that surviving megafauna on all continents—but particularly the ones with no hominin history—are mostly ones that are fast on their feet when escaping from predators and ones that have short gestation periods. This makes perfect sense, as nimble animals that can make a quick escape would fare better against a pack of humans armed with sharp weapons than large, slow animals that prefer to stand their ground in the face of a threat. Additionally, having more offspring would better help animals replenish unprecedented losses in the face of a new and especially lethal predator.
A recent study indicates that extinction likelihood was correlated with a multitude of traits such as island endemism, larger size, flat-footedness, and greater genetic distance to Old World tropical species30. As humans, we do not tend to think in exponential terms. But it is possible that extinct megafauna were not just somewhat more vulnerable to humans, but substantially more so—perhaps by orders of magnitude. This would make sense; even among extant megafauna, we see an enormous range in terms of ability to withstand human pressures. Take antelopes versus rhinos, for example.
The most obvious reason why anatomically modern humans (AMH) were so destructive ecologically is because they presented a novel threat to animal communities outside of Africa. Animals—like those of the Americas, Australasia, and islands—that had not evolved alongside hominins simply had no adaptations to hominin-specific hunting behavior. In Eurasia, a long coexistence period with non-AMH hominins meant the effect of AMH expansion on animal communities was not as severe as in other parts of the world. However, extinctions were still worse in Eurasia than in Africa which may at least partially be due to AMH being more lethal than pre-existing hominins; the fact that AMH rapidly replaced Neanderthals and Denisovans may lend support for this notion.

Humans presented a threat that could not be matched by any other predators. They were capable of bringing down animals of all sizes and age groups, which is highly detrimental to slow-reproducing megaherbivores as their survival depends on not having to face predation pressure during their prime reproductive years. They could also easily hunt megafaunal species to extinction and not doom themselves to starvation because they were capable of exploiting a wide variety of food sources, including small terrestrial animals, birds, fish, and plants.
It should be noted that we do not know the precise mechanisms involved in human-mediated extinction; it is quite likely that they were not accomplished solely through overhunting but also through second-order effects of overhunting such as ecosystem collapses through losses of keystone species such as megaherbivores, who were known to maintain vegetative openness via disturbance and act as seed and nutrient dispersers31 32. An excessive use of fire by humans is also speculated to have had effects33.
These would have affected not only megafauna but small animals as well, potentially explaining a number of non-megafaunal extinctions. There are countless other possibilities in this regard; could it also be, for instance, that humans menacing animals at grassy openings within forests or at mineral licks created an “atmosphere of fear” which caused them to avoid these essential locations, leading to deaths from malnutrition?
Moreover, humans eventually occupied essentially all environments, meaning there was no proper safe haven from human pressure the way there was from climate change (as refugia still existed during periods of climate stress). Some areas such as steep mountains, rainforests, and wetlands did offer some refuge for animals escaping predation pressure, but it was limited as humans were still present, just in lower numbers. And unlike climate change which could at times be a benefit, human presence ranged from neutral at best to decidedly destructive at worst for animal communities.
This is why some form of extinction inevitably follows permanent, widespread human settlement of new landmasses, regardless of whether the climate is changing profoundly or not. On islands, extinctions were very rapid and involved tiny numbers of humans. The Maori drove the Moa extinct within two centuries of arrival, despite the Maori population most likely never reaching more than 2,000 people during that period which also calls into question the idea that very low densities of humans cannot drive extinctions34. Non-megafaunal extinctions were numerous on New Zealand too35, just as they were on the Pacific island of Tonga36.
The arguments for human potency are compelling, but this article is not complete without tackling a common point raised against anthropogenic causation.
What About the Kill Sites?
An enduring objection to the idea that humans were the biggest culprit in the extinctions of megafauna globally is that there is not enough evidence of human predation on megafauna in the form of remains at “kill sites” which show clear evidence of hunting and butchery37 38. The logic initially seems sound—after all, should there not be abundant evidence if humans were indeed responsible for overhunting animals to such a massive extent?
The problem is that the expectation of abundant kill sites is an unreasonable one to begin with. The overwhelming majority of animals that ever live do not fossilize39, and the great lengths of time passed since extinction events further erodes evidence of human-megafauna interactions40. We have hundreds or even thousands of remains for some species or genera, but this is a very tiny amount when considering that there may have been millions of individuals living at any given time.
Glyptodonts were a clade of massive, hulking armadillo relatives with thick shells. They originated in South America and then spread to Central and southern parts of North America following the Great American Biotic Interchange. They have been around for tens of millions of years. Given their widespread range, it is easy to speculate that there could have been well over a million glyptodont individuals living at any given time. Despite this, as well as their massive size and peculiar anatomy and very ancient origins, we only have a few thousand preserved remains of them. If even these widespread, ancient, and hulking creatures are not preserved in high proportions, this means that fossilization must be an extremely rare phenomenon.

Some Pleistocene species and genera were not even known to us until the last few decades; others were thought to have gone extinct hundreds of thousands or millions of years ago only for a lucky find in the last few years to reveal that they made it into the Late Pleistocene. For example, the sabre-toothed cat Homotherium latidens was thought to have not survived into the Late Pleistocene of Europe only for a mandible to be discovered in the North Sea in 2009 dating to around 28 thousand radiocarbon years before present, hundreds of thousands of years younger than previously known Homotherium remains from that continent41. How many thousands of Homotherium latidens must have lived in Europe between those two finds, leaving no trace behind?
This also applies to humans. By now, it is increasingly accepted that humans were in the Americas prior to the Clovis era42 43. A total of perhaps hundreds of thousands, possibly even more, humans could have lived and died in the Americas during the Clovis period and earlier; yet, we only have a handful of Paleo-Indian remains and all of them only date to the Clovis and later (Arlington Springs Man, Luzia Woman, Buhl Woman, Anzick-1, etc.). Clearly, the animal and human specimens that actually fossilized and were found only represent a very tiny fraction of those that were present during a given period of time. Therefore, to expect there to be an abundance of animal remains from a select (usually short) period of time which just happen to show evidence of human processing is a big ask, especially when a large portion of those remains are fragmentary to begin with.
Over time, the physical traces of human hunting tend to disappear. In New Zealand, for instance, the extinction of the moa about 700 years ago is well-documented, with abundant evidence of overexploitation34 44. In the Caribbean islands by contrast—where the mid-Holocene extinction of ground sloths and monkeys is undeniably linked to humans—there is very little direct evidence of predation by the Amerindian settlers as indicated by a 2020 study45. The authors of the study attempt to reconcile the disappearance of the native fauna with the lack of kill sites by arguing that habitat alteration by humans may have been the primary driver of the losses rather than overhunting.
Yet, this defies all common sense; humans invariably heavily exploit naive fauna wherever they go. It is much more likely that evidence of human predation was preserved in New Zealand but not in the Caribbean islands simply due to differences in climate (temperate vs tropical) and in the length of time that has passed since both events (700 vs 5,000 years). The second has important ramifications: if the physical evidence for human predation is already so diminished after just a few thousand years, it stands to reason that it would be even rarer proportionally in extinctions dating back 12,000-45,000 years, as in the case of the continents.
Moreover, we have to ask how relevant the number of kill sites even is to the debate. Two powerful studies came out in the previous few years: one uses isotopes to show that mammoth was a major component of the diet of western Clovis46, and another found blood residues on Paleo-Indian blades that match those of proboscideans and equids (which are now extinct in the Americas)47. There is certainly no question that extinct megafauna were on the menu for early humans, meaning we have to question the logic of using the abundance—or lack thereof—of their butchered remains as an indicator of how much humans actually preyed on them.

But there is still one last issue to address with regard to the Late Quaternary Extinctions, one which has added to the difficulty of deciphering them.
Timing of Extinction in Relation to Human Arrival
One of the biggest hurdles in resolving extinction dynamics is disagreement over when humans first arrived on the continents. I go into detail on the controversies surrounding settlement dates and provide what I believe to be the most likely scenarios for the New World and Australia, but I will include a brief overview here as well. Although archeology has come a long way since its inception, accurately dating human remains and human-associated artefacts remains a persistent challenge. In many cases, it is not even certain whether some purported evidence of human occupation genuinely reflects human presence at all. As such, debates rage over the timing of human arrival on all of the continents. There is even disagreement over some islands like Madagascar48 which were settled much later and should therefore theoretically have a clearer record.
Notably, analysis of fossilized human footprints at White Sands National Park in New Mexico returned very old dates which sparked speculation about human presence in North America during or even prior to the Last Glacial Maximum between 25 and 19 thousand years ago49, and Madjedbebe site in northern Australia has been claimed to be as old as 65 kya50. The dates for White Sands and Madjedbebe are considerably older than the confirmed oldest sites in the Americas and Sahul respectively, and if they are accurate and represent a genuine widespread presence of humans in those regions so early on, can raise doubts about the importance of anthropogenic causation in extinctions there. Still, there is controversy regarding those dates. Madjedbebe in particular is highly contentious51. Some are not convinced by the evidence for the White Sands footprints being so old either52, but there appears to be relatively more acceptance for it in the scientific community than for Madjedbebe.

However, genetics can come to the rescue in times like these as analysis of modern and ancient human individuals can tell us a great deal about when their ancestors must have arrived. Archeology is not my strong suit, so I will refrain from analyzing the veracity of the dating methods used at White Sands. But the genetic evidence indicates that Paleo-Indians, the ancestors of modern Native Americans, arrived in the Americas south of the erstwhile ice sheet around 16 thousand years ago42.
If there were truly people present in the White Sands area during the LGM, the total lack of human remains until 13 kya and the absence of other fully verified archeological sites prior to 16 thousand years ago in North America suggests that they likely represent some sort of short-lived, small, geographically restricted population that may have even been a genetic dead. Hence, it is not necessary that whatever interactions that the purported White Sands population had with megafauna were relevant or meaningful in the grand scheme of things. As for Australian Aboriginals, genetic analysis points to an arrival in Sahul shortly after 50 thousand years ago53.
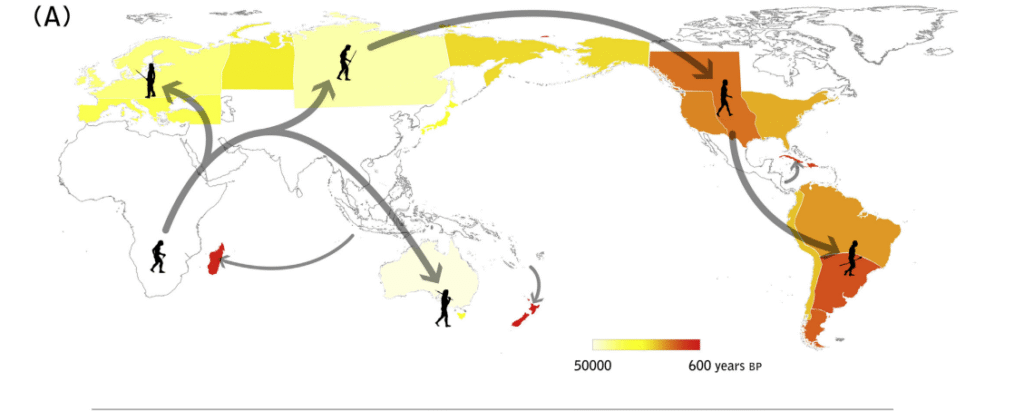
A 16 kya arrival for Paleo-Indians clearly precedes the emergence of the Clovis culture in North America and the extinctions in the New World by around 3 thousand years. Some may argue that what appears to be a few millennia-long “coexistence” between humans and megafauna rather than immediate megafaunal collapse refutes the idea of mainly human-driven extinctions and warrants some sort of significant climatic input5 54, but in fact this is perfectly congruent with that scenario. The initial population of humans arriving on any landmass is always small and reaching an appreciable population density on continents will be a lengthy process given the vast areas that need to be settled; this would have been especially true for the Americas, as North and South America were colonized almost simultaneously by the same small founding population. This meant a long process of exploring, adapting to, and gradually populating two immense continents with ferocious predators and unfamiliar landscapes.

The Clovis may not have been the first people in the Americas but the period starting around 13 kya is still significant in that it may represent the first time that humans in North and South America achieved a population density large enough to meaningfully impact megafauna. Indeed, this is when remains of humans first start to appear in the fossil record. This population growth would have been augmented with the concurrent adoption of Clovis and Fishtail blades which were ideal for hunting big-game12 55 with the result that once extinctions began, they concluded rapidly. Like most things in life, this was not a linear process.
It makes sense that in both the New World and Australia, extinction onset lagged initial human arrival by a few millennia. It is quite possible that during the initial phase of colonization, people were so rare on the landscape that the vast majority of megafaunal individuals went their entire lives without ever seeing a person. The fact that extinctions were not instantaneous there as they were on islands should not be used as proof that humans alone were not capable of driving continent-wide extinctions, as the length of time needed for human settlers to become a lethal force would have been far greater on said continents compared to islands which were far smaller and had even more naive megafauna.
Conclusion and Sequel
While I had long dreamed of starting a prehistory-themed blog, I did not envision this topic to become so prominent within it. However, reading about Pleistocene paleoecology inevitably means running into mentions of these extinctions as well as, unfortunately, a seemingly endless amount of questionable analysis on their causes as well. I must admit that as a lifelong lover of nature and history, this topic has become somewhat personal for me. And as someone generally invested in the truth and accuracy, I felt a strong need to offer scrutiny and pushback against dubious narratives as well as cut through a lot of the confusion I have seen.
On one hand, it appears (at least anecdotally) that the number of experts who believe in a mainly or exclusively climatic cause for the global LQE has been dwindling; on the other, there is no shortage of people who continue to downplay the human role and exaggerate the importance of climate. Proponents of these viewpoints tend to portray their stances as being nuanced in contrast to more human-centric explanations. But as shown in the first article of this series, true nuance with regard to the topic would involve appreciating the complexities of animal responses to climate change as well as the complex effects of climate change itself, which in turn would lead one to question the idea that climate could have been nearly as impactful as the ascent of man.
In the previous article, we showed that due to the nature of Earth’s climate system, geography, and animal communities, the effects of climate change at the individual species level do not necessarily translate to effects at the community level, nor are its effects at the regional scale necessarily relevant at the continental scale. There is no denying that some animals may have been struggling during any given climate phase of the Late Quaternary; however, it is not possible that all or most species were simultaneously on the brink during said phases. It is even more difficult to believe that such a thing would coincidentally happen shortly following human arrival, as is often claimed.
We started this article by concluding arguments against climate theories by drawing attention to extinction patterns along biogeographic lines, showing that extinction was worst in the continents where native megafauna were most vulnerable to human pressure as opposed to those where climate change struck the hardest. It is impossible to claim that the contribution of climate change to extinctions was comparable to or greater than that of humans (or that the two causes were synergistic), and then come up empty-handed when trying to establish a link between extinction severity and climate change magnitude by continent.
We then analyzed the human dimension, exploring why it makes perfect sense that humans were so lethal to animal communities, how humans may have settled various continents, and finally what the alleged lack of megafaunal “kill sites” actually means. The key takeaways are:
- Humans were coming up against animals which had no defenses towards hominin predation whatsoever (in the New World, Australia, and islands) or those which had not developed defenses against Homo sapiens specifically (as in Eurasia).
- Humans were versatile and could exploit a wide variety of game, and move on to plant and small animal food sources after having exhausted the supply of larger animals. They were far more destructive than any apex predator as a result.
- Unlike climate change, humans occupied more or less all environments, meaning there was no escape from them.
- Mechanisms beyond plain overhunting may have been involved in extinctions, such as habitat alteration through the loss of keystone predators and/or excessive use of fire, among others.
- There should not be an expectation of rapid extinctions following human arrival on any given continent, as it should take a long time for humans to spread out and reach an appreciable population density on large landmasses.
- The relative paucity of direct evidence for human predation on extinct megafauna cannot be used as an argument against mostly human-driven extinctions, as fossilization is inherently very rare and extinction windows were often short.
Some people may continue to object to the notion of majority human driven extinctions on the basis of human population densities allegedly being too low and a perceived lack of evidence for human predation on megafauna such as kill sites. However, this is epistemologically backwards. If the evidence we have clearly indicates extinctions shortly following human arrival combined with an inadequacy of climatic causes to properly explain those extinctions, then we should question our assumptions about the importance of high population density and/or the abundance of kill sites, rather than whether or not humans played the major role. The picture of small populations of early humans wiping out large numbers of megafauna and then leaving little direct evidence of interaction is admittedly shocking and hard to imagine, but it is most certainly not impossible.
We may always be evaluating the precise importance of climate change in these extinctions, and we may never truly know what it was. There is decent reason to believe it was significant in some regions. But regarding the question of whether or not climate was roughly as or more impactful than human activity on the global scale, there is simply no more room for debate. The answer, based on all the available evidence, is a resounding no.
In spite of the foundations for belief in climate-heavy theories for LQE being remarkably weak, there is no shortage of studies that argue for a much larger impact from climate change than humans in various parts of the world. What do we make of this? Well, the simple answer is that these studies are flawed, in some cases profoundly so. The next part of this series will take a look at a selection of these studies to demonstrate why we should maintain skepticism toward notions of climate-centric causes for extinction even in the face of their popularity. We will also be exploring the role that bias may be playing in the extinction debate and why it is important to come to terms with the reality revealed by the evidence.
Part 3: Bad Science and Facing Reality
References
- Sandom, C., Faurby, S., Sandel, B., & Svenning, J.-C. (2014). Global late Quaternary megafauna extinctions linked to humans, not climate change. Proceedings of the Royal Society B: Biological Sciences, 281(1787), 20133254. https://doi.org/10.1098/rspb.2013.3254[↩][↩][↩][↩]
- Araujo, B. B. A., Oliveira-Santos, L. G. R., Lima-Ribeiro, M. S., Diniz-Filho, J. A. F., & Fernandez, F. A. S. (2017). Bigger kill than chill: The uneven roles of humans and climate on late Quaternary megafaunal extinctions. Quaternary International, 431, 216–222. https://doi.org/10.1016/j.quaint.2015.10.045[↩][↩]
- Lemoine, R. T., Buitenwerf, R., & Svenning, J.-C. (2023). Megafauna extinctions in the late-Quaternary are linked to human range expansion, not climate change. Anthropocene, 44, 100403. https://doi.org/10.1016/j.ancene.2023.100403[↩]
- Meltzer, D. J. (2020). Overkill, glacial history, and the extinction of North America’s Ice Age megafauna. Proceedings of the National Academy of Sciences, 117(46), 28555–28563. https://doi.org/10.1073/pnas.2015032117[↩]
- Cione, A. L., Tonni, E. P., & Soibelzon, L. (2009). Did humans cause the Late Pleistocene-Early Holocene mammalian extinctions in South America in a context of shrinking open areas?. American megafaunal extinctions at the end of the Pleistocene, 125-144.[↩][↩]
- Wroe, S., Field, J. H., Archer, M., Grayson, D. K., Price, G. J., Louys, J., Faith, J. T., Webb, G. E., Davidson, I., & Mooney, S. D. (2013). Climate change frames debate over the extinction of megafauna in Sahul (Pleistocene Australia-New Guinea). Proceedings of the National Academy of Sciences, 110(22), 8777–8781. https://doi.org/10.1073/pnas.1302698110[↩]
- Koch, P. L., & Barnosky, A. D. (2006). Late Quaternary Extinctions: State of the Debate. Annual Review of Ecology, Evolution, and Systematics, 37(1), 215–250. https://doi.org/10.1146/annurev.ecolsys.34.011802.132415[↩]
- Surovell, T. A., Pelton, S. R., Anderson-Sprecher, R., & Myers, A. D. (2015). Test of Martin’s overkill hypothesis using radiocarbon dates on extinct megafauna. Proceedings of the National Academy of Sciences, 113(4), 886–891. https://doi.org/10.1073/pnas.1504020112[↩]
- Pires, M. M., Koch, P. L., Fariña, R. A., de Aguiar, M. A. M., dos Reis, S. F., & Guimarães, P. R. (2015). Pleistocene megafaunal interaction networks became more vulnerable after human arrival. Proceedings of the Royal Society B: Biological Sciences, 282(1814), 20151367. https://doi.org/10.1098/rspb.2015.1367[↩]
- Bampi, H., Barberi, M., & Lima-Ribeiro, M. S. (2022). Megafauna kill sites in South America: A critical review. Quaternary Science Reviews, 298, 107851. https://doi.org/10.1016/j.quascirev.2022.107851[↩]
- Politis, G. G., Messineo, P. G., Stafford, T. W., & Lindsey, E. L. (2019). Campo Laborde: A Late Pleistocene giant ground sloth kill and butchering site in the Pampas. Science Advances, 5(3), eaau4546. https://doi.org/10.1126/sciadv.aau4546[↩]
- Prates, L., & Perez, S. I. (2021). Late Pleistocene South American megafaunal extinctions associated with rise of Fishtail points and human population. Nature Communications, 12(1). https://doi.org/10.1038/s41467-021-22506-4[↩][↩]
- Prates, L., Medina, M. E., & Perez, S. I. (2025). Extinct megafauna dominated human subsistence in southern South America before 11,600 years ago. Science Advances, 11(40). https://doi.org/10.1126/sciadv.adx2615[↩]
- Williams, J. W., Shuman, B. N., & III, T. W. (2001). Dissimilarity Analyses of Late-Quarternary Vegetation and Climate in Eastern North America. Ecology, 82(12), 3346. https://doi.org/10.2307/2680157[↩]
- Saltré, F., Rodríguez-Rey, M., Brook, B. W., Johnson, C. N., Turney, C. S. M., Alroy, J., Cooper, A., Beeton, N., Bird, M. I., Fordham, D. A., Gillespie, R., Herrando-Pérez, S., Jacobs, Z., Miller, G. H., Nogués-Bravo, D., Prideaux, G. J., Roberts, R. G., & Bradshaw, C. J. A. (2016). Climate change not to blame for late Quaternary megafauna extinctions in Australia. Nature Communications, 7(1). https://doi.org/10.1038/ncomms10511[↩]
- van der Kaars, S., Miller, G. H., Turney, C. S. M., Cook, E. J., Nürnberg, D., Schönfeld, J., Kershaw, A. P., & Lehman, S. J. (2017). Humans rather than climate the primary cause of Pleistocene megafaunal extinction in Australia. Nature Communications, 8(1). https://doi.org/10.1038/ncomms14142[↩]
- Barnosky, A. D., & Lindsey, E. L. (2010). Timing of Quaternary megafaunal extinction in South America in relation to human arrival and climate change. Quaternary International, 217(1-2), 10–29. https://doi.org/10.1016/j.quaint.2009.11.017[↩]
- Broughton, J. M., & Weitzel, E. M. (2018). Population reconstructions for humans and megafauna suggest mixed causes for North American Pleistocene extinctions. Nature Communications, 9(1). https://doi.org/10.1038/s41467-018-07897-1[↩]
- Faurby, S., & Svenning, J.-C. (2016). The asymmetry in the Great American Biotic Interchange in mammals is consistent with differential susceptibility to mammalian predation. Global Ecology and Biogeography, 25(12), 1443–1453. https://doi.org/10.1111/geb.12504[↩][↩]
- Stewart, M., Peters, C., Ziegler, M. J., Carleton, W. C., Roberts, P., Boivin, N., & Groucutt, H. S. (2025). The state of the late Quaternary megafauna extinction debate: a systematic review and analysis. Frontiers in Mammal Science, 4. https://doi.org/10.3389/fmamm.2025.1678231[↩]
- Stuart, A. J. (2014). Late Quaternary megafaunal extinctions on the continents: a short review. Geological Journal, 50(3), 338–363. https://doi.org/10.1002/gj.2633[↩][↩][↩]
- Stuart, A. J., & Lister, A. M. (2007). Patterns of Late Quaternary megafaunal extinctions in Europe and northern Asia.[↩]
- Wright, H. E. (1977). Quaternary Vegetation History-Some Comparisons Between Europe and America. Annual Review of Earth and Planetary Sciences, 5(1), 123–158. https://doi.org/10.1146/annurev.ea.05.050177.001011[↩]
- Binney, H., Edwards, M., Macias-Fauria, M., Lozhkin, A., Anderson, P., Kaplan, J. O., Andreev, A., Bezrukova, E., Blyakharchuk, T., Jankovska, V., Khazina, I., Krivonogov, S., Kremenetski, K., Nield, J., Novenko, E., Ryabogina, N., Solovieva, N., Willis, K., & Zernitskaya, V. (2017). Vegetation of Eurasia from the last glacial maximum to present: Key biogeographic patterns. Quaternary Science Reviews, 157, 80–97. https://doi.org/10.1016/j.quascirev.2016.11.022[↩]
- Tallavaara, M., Luoto, M., Korhonen, N., Järvinen, H., & Seppä, H. (2015). Human population dynamics in Europe over the Last Glacial Maximum. Proceedings of the National Academy of Sciences, 112(27), 8232–8237. https://doi.org/10.1073/pnas.1503784112[↩]
- Goebel, T. (1999). Pleistocene human colonization of Siberia and peopling of the Americas: An ecological approach. Evolutionary Anthropology: Issues, News, and Reviews, 8(6), 208–227. https://doi.org/10.1002/(sici)1520-6505(1999)8:6%3C208::aid-evan2%3E3.0.co;2-m[↩]
- Svenning, J.-C., Lemoine, R. T., Bergman, J., Buitenwerf, R., Roux, E. L., Lundgren, E., Mungi, N., & Pedersen, R. Ø. (2024). The late-Quaternary megafauna extinctions: Patterns, causes, ecological consequences and implications for ecosystem management in the Anthropocene. Cambridge Prisms: Extinction, 2, e5. https://doi.org/10.1017/ext.2024.4[↩]
- Helm, C. W., Carr, A. S., Cawthra, H. C., De, J. C., Dixon, M. G., Pieter-Jan Gräbe, Guy, & Venter, J. A. (2023). Tracking the extinct giant Cape zebra (Equus capensis) on the Cape south coast of South Africa. Quaternary Research, 114, 178–190. https://doi.org/10.1017/qua.2023.1[↩]
- DeMenocal, P. B., & Tierney, J. E. (2012). Green Sahara: African humid periods paced by Earth’s orbital changes. Nature Education Knowledge, 3(10), 12.[↩]
- Lemoine, R. T., Buitenwerf, R., Sören Faurby, & Jens‐Christian Svenning. (2025). Phylogenetic Evidence Supports the Effect of Traits on Late‐Quaternary Megafauna Extinction in the Context of Human Activity. Global Ecology and Biogeography, 34(7). https://doi.org/10.1111/geb.70078[↩]
- Owen-Smith, N. (1987). Pleistocene extinctions: the pivotal role of megaherbivores. Paleobiology, 13(3), 351–362. https://doi.org/10.1017/s0094837300008927[↩]
- Smith, F. A., Elliott Smith, E. A., Hedberg, C. P., Lyons, S. K., Pardi, M. I., & Tomé, C. P. (2023). After the mammoths: the ecological legacy of late Pleistocene megafauna extinctions. Cambridge Prisms: Extinction, 1(9), 1–55. https://doi.org/10.1017/ext.2023.6[↩]
- Miller, G. H. (2005). Ecosystem Collapse in Pleistocene Australia and a Human Role in Megafaunal Extinction. Science, 309(5732), 287–290. https://doi.org/10.1126/science.1111288[↩]
- Holdaway, R. N., Allentoft, M. E., Jacomb, C., Oskam, C. L., Beavan, N. R., & Bunce, M. (2014). An extremely low-density human population exterminated New Zealand moa. Nature Communications, 5(1). https://doi.org/10.1038/ncomms6436[↩][↩]
- Holdaway, R. N., Worthy, T. H., & Tennyson, A. J. D. (2001). A working list of breeding bird species of the New Zealand region at first human contact. New Zealand Journal of Zoology, 28(2), 119–187. https://doi.org/10.1080/03014223.2001.9518262[↩]
- Steadman, D. W., Pregill, G. K., & Burley, D. V. (2002). Rapid prehistoric extinction of iguanas and birds in Polynesia. Proceedings of the National Academy of Sciences of the United States of America, 99(6), 3673–3677. https://doi.org/10.1073/pnas.072079299[↩]
- Grayson, D. K., & Meltzer, D. J. (2003). A requiem for North American overkill. Journal of Archaeological Science, 30(5), 585–593. https://doi.org/10.1016/s0305-4403(02)00205-4[↩]
- Field, J., Wroe, S., Trueman, C. N., Garvey, J., & Wyatt-Spratt, S. (2013). Looking for the archaeological signature in Australian Megafaunal extinctions. Quaternary International, 285, 76–88. https://doi.org/10.1016/j.quaint.2011.04.013[↩]
- Fiedel, S., & Haynes, G. (2004). A premature burial: comments on Grayson and Meltzer’s “Requiem for overkill.” Journal of Archaeological Science, 31(1), 121–131. https://doi.org/10.1016/j.jas.2003.06.004[↩]
- Surovell, T. A., & Grund, B. S. (2012). The Associational Critique of Quaternary Overkill and why it is Largely Irrelevant to the Extinction Debate. American Antiquity, 77(4), 672–687. https://doi.org/10.7183/0002-7316.77.4.672[↩]
- Reumer, J. W. F., Rook, L., Van Der Borg, K., Post, K., Mol, D., & De Vos, J. (2003). Late Pleistocene survival of the saber-toothed cat Homotherium in northwestern Europe. Journal of Vertebrate Paleontology, 23(1), 260–262.[↩]
- Llamas, B., Fehren-Schmitz, L., Valverde, G., Soubrier, J., Mallick, S., Rohland, N., Nordenfelt, S., Valdiosera, C., Richards, S. M., Rohrlach, A., Romero, M. I. B., Espinoza, I. F., Cagigao, E. T., Jiménez, L. W., Makowski, K., Reyna, I. S. L., Lory, J. M., Torrez, J. A. B., Rivera, M. A., & Burger, R. L. (2016). Ancient mitochondrial DNA provides high-resolution time scale of the peopling of the Americas. Science Advances, 2(4), e1501385. https://doi.org/10.1126/sciadv.1501385[↩][↩]
- Waters, M. R., Keene, J. L., Forman, S. L., Prewitt, E. R., Carlson, D. L., & Wiederhold, J. E. (2018). Pre-Clovis projectile points at the Debra L. Friedkin site, Texas—Implications for the Late Pleistocene peopling of the Americas. Science Advances, 4(10), eaat4505. https://doi.org/10.1126/sciadv.aat4505[↩]
- Oskam, C. L., Allentoft, M. E., Walter, R., Scofield, R. P., Haile, J., Holdaway, R. N., Bunce, M., & Jacomb, C. (2012). Ancient DNA analyses of early archaeological sites in New Zealand reveal extreme exploitation of moa (Aves: Dinornithiformes) at all life stages. Quaternary Science Reviews, 52, 41–48. https://doi.org/10.1016/j.quascirev.2012.07.007[↩]
- Orihuela, J., Viñola, L. W., Jiménez Vázquez, O., Mychajliw, A. M., Hernández de Lara, O., Lorenzo, L., & Soto-Centeno, J. A. (2020). Assessing the role of humans in Greater Antillean land vertebrate extinctions: New insights from Cuba. Quaternary Science Reviews, 249, 106597. https://doi.org/10.1016/j.quascirev.2020.106597[↩]
- Chatters, J. C., Potter, B. A., Fiedel, S. J., Morrow, J. E., Jass, C. N., & Wooller, M. J. (2024). Mammoth featured heavily in Western Clovis diet. Science Advances, 10(49). https://doi.org/10.1126/sciadv.adr3814[↩]
- Moore, C., Kimball, L. R., Goodyear, A. C., Brooks, M. J., I. Randolph Daniel, West, A., Taylor, S., Weber, K., Fagan, J., & Walker, C. (2023). Paleoamerican exploitation of extinct megafauna revealed through immunological blood residue and microwear analysis, North and South Carolina, USA. Scientific Reports, 13(1). https://doi.org/10.1038/s41598-023-36617-z[↩]
- Mitchell, P. (2019). Settling Madagascar: When Did People First Colonize the World’s Largest Island? The Journal of Island and Coastal Archaeology, 1–20. https://doi.org/10.1080/15564894.2019.1582567[↩]
- Bennett, M. R., Bustos, D., Pigati, J. S., Springer, K. B., Urban, T. M., Holliday, V. T., Reynolds, S. C., Budka, M., Honke, J. S., Hudson, A. M., Fenerty, B., Connelly, C., Martinez, P. J., Santucci, V. L., & Odess, D. (2021). Evidence of humans in North America during the Last Glacial Maximum. Science, 373(6562), 1528–1531. https://doi.org/10.1126/science.abg7586[↩]
- Clarkson, C. (2017). Human occupation of northern Australia by 65,000 years ago. Nature, 547(7663), 306–310. https://doi.org/10.1038/nature22968[↩]
- O’Connell, J. F., Allen, J., Williams, M. A. J., Williams, A. N., Turney, C. S. M., Spooner, N. A., Kamminga, J., Brown, G., & Cooper, A. (2018). When did Homo sapiens first reach Southeast Asia and Sahul? Proceedings of the National Academy of Sciences, 115(34), 8482–8490. https://doi.org/10.1073/pnas.1808385115[↩]
- Oviatt, C. G., Madsen, D. B., Rhode, D., & Davis, L. G. (2022). A critical assessment of claims that human footprints in the Lake Otero basin, New Mexico date to the Last Glacial Maximum. Quaternary Research, 111, 138–147. https://doi.org/10.1017/qua.2022.38[↩]
- Tobler, R., Rohrlach, A., Soubrier, J., Bover, P., Llamas, B., Tuke, J., Bean, N., Abdullah-Highfold, A., Agius, S., O’Donoghue, A., O’Loughlin, I., Sutton, P., Zilio, F., Walshe, K., Williams, A. N., Turney, C. S. M., Williams, M., Richards, S. M., Mitchell, R. J., & Kowal, E. (2017). Aboriginal mitogenomes reveal 50,000 years of regionalism in Australia. Nature, 544(7649), 180–184. https://doi.org/10.1038/nature21416[↩]
- Metcalf, J. L., Turney, C., Barnett, R., Martin, F., Bray, S. C., Vilstrup, J. T., Orlando, L., Salas-Gismondi, R., Loponte, D., Medina, M., De Nigris, M., Civalero, T., Fernández, P. M., Gasco, A., Duran, V., Seymour, K. L., Otaola, C., Gil, A., Paunero, R., & Prevosti, F. J. (2016). Synergistic roles of climate warming and human occupation in Patagonian megafaunal extinctions during the Last Deglaciation. Science Advances, 2(6), e1501682. https://doi.org/10.1126/sciadv.1501682[↩]
- Kilby, J. D., Surovell, T. A., Huckell, B. B., Ringstaff, C. W., Hamilton, M. J., & Haynes, C. V. (2022). Evidence supports the efficacy of Clovis points for hunting proboscideans. Journal of Archaeological Science: Reports, 45, 103600. https://doi.org/10.1016/j.jasrep.2022.103600[↩]
Excellent work with sharp arguments and string logic. You made your case in a convincing way. Of course, only the novelty of the modern human hyper- predator could have caused such a bizarre extinction pattern as is the subject here. I also follow the debate since a while, but you brought some new thoughts into it making it even harder to believe that climate has anything to do with the loss of our megafauna..
Thank you for the kind words, Matthias.