There are few places on the planet as distinctive as Australia. With its isolated geographical location and iconic landscapes, it captures the imagination of people the world over. Picturesque red deserts, glistening beaches, and Eucalyptus woodlands may provide its most characteristic scenery, but arcane rainforests, snow-covered mountains, and much more can be found on the continent as well.



As fascinating as today’s Australian fauna are, something is missing. Few of them classify as megafauna, or animals larger than 45 kg. Up to 23 out of 24 genera of Australian land animals weighing more than 45kg disappeared towards the end of the Late Quaternary, mostly by 40 thousand years ago1. Among the genera that were lost included the largest known marsupial, a giant flightless bird, a massive relative of the Komodo dragon, a marsupial version of the lion, and a terrestrial crocodile. It is hard to imagine the continent once hosted such beasts, given that the largest land animal there today is the red kangaroo.
What happened to all these enigmatic creatures? The Australian extinctions serve as both an extension of and a focal point within the broader debate surrounding the disappearance of most of the world’s megafauna during the Late Pleistocene across multiple continents, with humans and climate change of course being the main suspects. Those who blame humans argue that overhunting and/or landscape modification were the root cause of extinction in Australia, while those who favor climatic explanations argue that it was instead increasing aridity that pushed the fauna of this already dry continent over the edge.
After having done extensive research on the topic, I will argue comprehensively for the idea that humans played the primary role in the disappearance of megafauna, focusing mainly on the following points:
- Aboriginal arrival closely coinciding with a clear extinction wave
- Serious flaws in climate arguments, including a lack of evidence for major climatic instability during the extinction window 50-40 thousand years ago (kya)
- High susceptibility of Australian megafauna to human predation
- Potential contribution of anthropogenic modification of Australian landscapes to extinctions
To some, it may seem audacious to suggest that climate played at most a minor part in the extinction of Australian megafauna. This post will therefore use extensive references and logical reasoning to argue for why it is difficult to conceive of plausible alternate explanations to mainly human causation. Uncertainties in the research will be made clear, as will my own personal speculations.
Before going further, I would like to note that the geographic terminology for this part of the world can get confusing. There are several names for broader regions that include Australia. For the purpose of reducing confusion, I will use Australia synonymously with mainland Australia while Australasia (though more of a biogeographic term) will be used to refer to the modern region encompassing Australia, New Guinea, and Tasmania. Sahul will refer to the prehistoric continent that united all three.
With that said, let us get to the bottom of this mystery in the land down under.
Background
To assess what happened to the megafauna, it may be best to set context by describing Australia on the eve of the extinctions. It was the first half of the period known as Marine Isotope Stage 3, an important phase of the last glaciation. It was cooler than today but not as cold as the Last Glacial Maximum (LGM) tens of thousands of years later. Like today, Australia hosted desert, savannah, dry scrub, swamp, and forest, although the exact geographic distributions of these biomes differed somewhat from their present ones.
Lower sea levels during glacials exposed continental shelves and created land bridges connecting mainland Australia to several islands in the Australasian realm, most notably New Guinea and Tasmania. A land bridge between Australia and New Guinea was present by the time humans arrived, while the land bridge between Australia and Tasmania formed slightly later. The combined landmass created a paleocontinent known as Sahul.
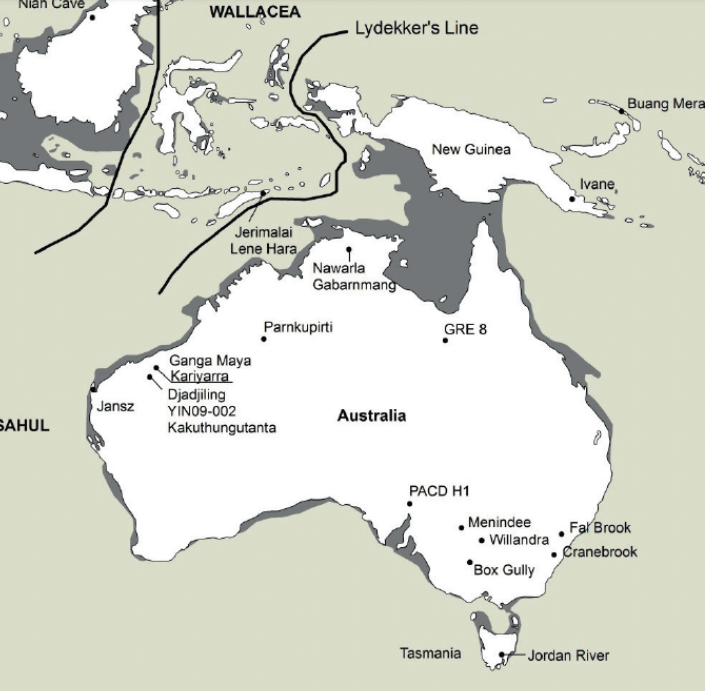
continental shelves in grey. Allen and O’Connell (2014). Click to expand
Extinct megafaunal species present at the time included Diprotodon optatum, a nearly 3-ton animal resembling a giant koala and perhaps the largest known marsupial; Genyornis newtoni, the last known member of a group of large, flightless Australian birds known as thunderbirds; and Thylacoleo carnifex, also called the marsupial lion. An unnamed species of Meiolania, a genus of large turtles widespread across Australasia, was present as well. It may have been the biggest of its genus at 2-3 meters long.

Thylacoleo (bottom center), and Varanus/Megalania (bottom left). Art by Peter Trusler
Quinkana fortirostrum was a terrestrial crocodile of the Mekosuchine clade. Varanus priscus, also known as Megalania, was a massive relative of the Komodo that may have been 5 meters long. Additionally, there were multiple species of large kangaroos belonging to the extinct genera Procoptodon (short faced kangaroos) and Protemnodon (giant kangaroos), while the extant Macropus genus (modern grey kangaroos) contained more members than the current two surviving species.



A biogeographic barrier known as the Wallace line separates Australasia from southeast Asia. As a result, Australasia has been isolated for tens of millions of years, which is why the overall composition of its flora and fauna diverged from other realms to such a significant degree. It is the only place where marsupials are the dominant form of mammals, and no placental land mammals besides bats and rodents were present before humans and the animals that humans introduced-like dingoes-arrived. This is important context when we get to discussing the fate of those fauna.
Now that we understand what kind of continent humans were about to discover, we can move on to pinpointing when they actually got there and the implications of it.
The Arrival
The trouble with tying human arrival to the megafaunal extinctions is that scientists are not fully confident as to when humans first arrived on any given continent. The information is important because conventional logic on this topic would suggest that extinction should immediately follow human entry. Time between arrival and extinction is not the end-all-be-all for determining human culpability here as there are many factors which can delay extinction at the hands of people, but it is useful to get an idea of when people arrived nonetheless. So how do we figure out when people actually got to Australia?
Since archeologists constantly debate the dates of very old archeological sites, insight from genetics-specifically concerning Papuans and indigenous Australians-is absolutely pivotal in solving this puzzle. We can start by noting that genetic research indicates Neanderthal ancestry introgressed into the common ancestors of all non-Africans in the Middle East around 54-49 kya2. Shortly after, the East Eurasian (also known as Eastern non-African) clade split from West Eurasians. Australasian populations fit within the East Eurasian clade, along with East and Southeast Asians while Europeans and West Asians fall within the West Eurasian clade.
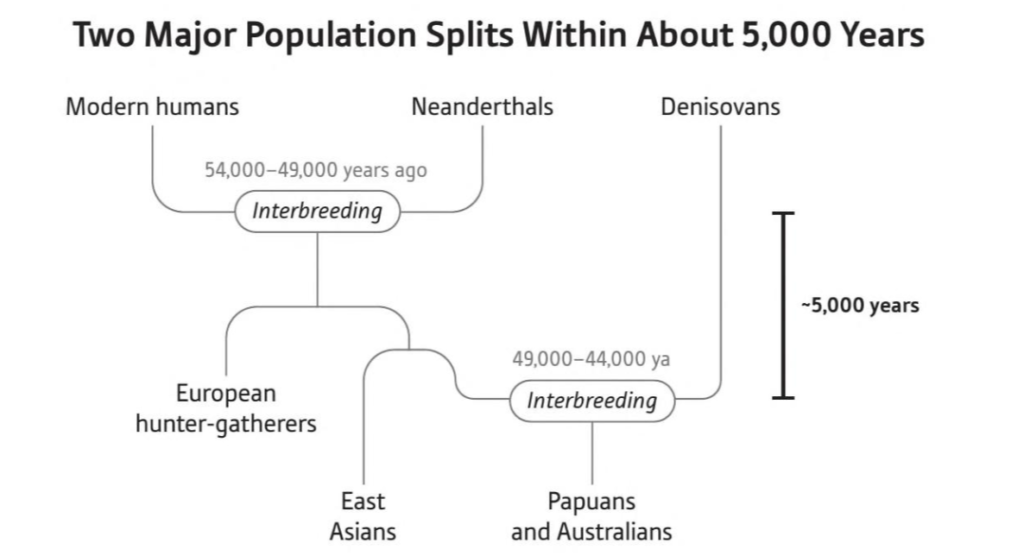
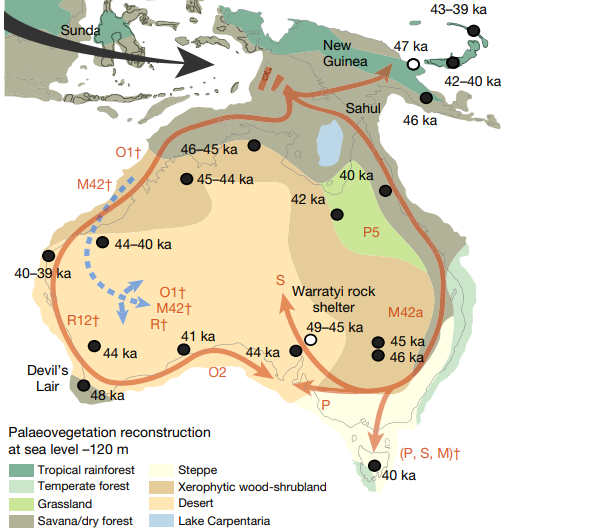
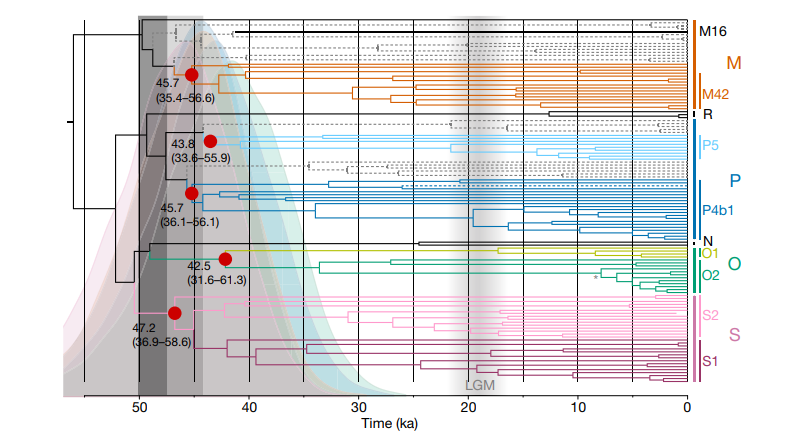
Next came the split of the ancestors of Australasians from the rest of the East Eurasian clade. After ~49 kya, the ancestors of Australasians absorbed a large amount of Denisovan ancestry, likely in southeast Asia. From there they went on to Sahul; a study on Aboriginal mitogenomes indicates a rapid colonization of Sahul from the northern part of the continent soon after 50 kya3. This aligns with the oldest reliable archeological sites in Australia which date to around 48-47 kya4 5.

These key events starting from Neanderthal admixture into the ancestors of non-Africans and ending with the settlement of Sahul happened in short succession. This was a rapid eastward migration across Eurasia and into Sahul during the Initial Upper Paleolithic, a period of technological and cultural leaps among humans. The main takeaway here is that Australia was settled slightly later than 50 kya, not earlier.
It is commonly claimed that people arrived in Australia around 65 kya, which if true could imply an extended coexistence between humans and megafauna. However, this figure is based solely on the controversial Madjedbebe site in northern Australia, whose contents are probably much younger5. There are no reliable dates for Homo sapiens presence this old in most of Eurasia, and this date does not align with what we know about Aboriginal genetics as outlined above.
Hypothetically speaking, if people were indeed present in Australia 65 kya+, the human group(s) present there at the time almost certainly were not the primary ancestors of modern Aboriginals. More importantly, the lack of associated archeological sites left behind by these alleged pre-Aboriginal arrivals would suggest they were probably too small in number to drastically impact their environment anyway. The implications are that we can largely ignore Madjedbebe and reasonably set the date for Aboriginal settlement to about 48-47 kya.
Pinpointing the Time of Extinctions
Now that we have a timeframe for when Aboriginals arrived, we need to determine when the extinctions (or at least most of them) took place. This is important to do for two reasons:
- Some argue that the extinctions were staggered and that there was no single brief period when Australian megafauna died off en masse6. Rather, increasingly intense climate cycles over the past few hundred thousand years wiped out megafauna in many waves such that there were hardly any present by the time humans arrived.
- Proponents of staggered extinctions also contend that those surviving megafauna then coexisted with humans for many thousands of years before disappearing, suggesting that humans were not responsible because they should have immediately caused extinction upon arrival as opposed to delaying it7.
In other words, the extinctions in Australia were not so much “Late Pleistocene” as they were “Pleistocene” since they were spread out over both the Middle and Late Pleistocene. We may refer to this as the “attrition hypothesis” for convenience. However, there are major issues with it as it is not clear at all that Middle Pleistocene species were mostly extinct by the time humans arrived. Evidence from places like Naracoorte cave indicates that many species indeed made it well into the Late Pleistocene8. The Signor-Lipps effect combined with an especially scanty fossil record for Australia is likely causing the extinctions to appear far more staggered than they really were. Towards the end of the next section, we will explain why it is unlikely that such high, staggered losses took place.
As for the second, there are major doubts about the validity of late-surviving megafauna in Australia, with outlying dates usually resulting from erroneous dating methods9. Further, a few late surviving outliers do not diminish the big picture of an extinction pulse taking place at a given time. In fact, due to the great size of Australia, it would be unsurprising if some stragglers did hang on for much longer than others in some locations even if animal communities were wiped out quickly overall.
Going back to the original question, can we determine when the megafauna, or at least most of them, vanished? Coprophilous fungi, or dung fungi, can give us major clues. Dung fungi are associated with the digestive process of herbivorous animals and can be used by scientists to track the abundance of megafauna at locations through time. Higher levels of dung fungi usually indicate higher animal density; a crash in levels with no recovery can indicate extinction. Though concerns exist on the reliability and utility of dung fungi as a proxy, taking certain precautions can resolve them10.
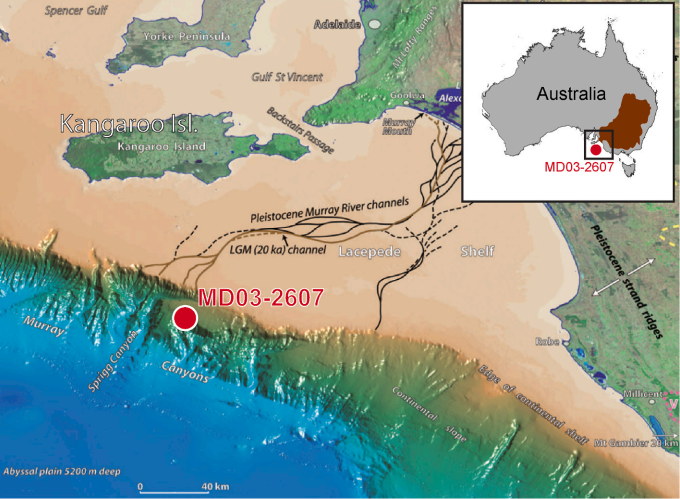
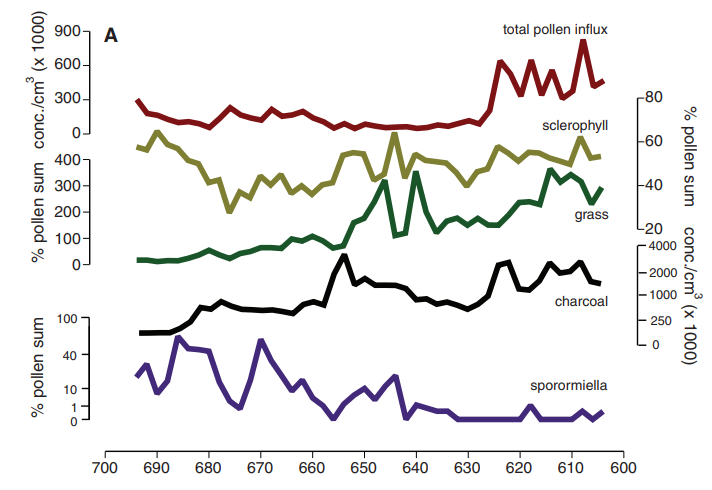
A number of studies using dung fungal analysis have been published for Australia. Johnson et al. 2015 reported a decline in dung fungi at Lynch’s crater in northeast Australia around 40 kya11. A 2017 study analyzed an ocean core off Cape Pasley in southwestern Australia while a 2023 study analyzed another near Kangaroo island (recording data from the Murray-Darling basin in interior southeastern Australia)12 13. Both ocean cores revealed that dung fungal spores plummeted to low levels around 43 kya. In Caledonia Fen, a high-elevation site near the coast of southeastern Australia, dung fungi plummeted somewhere between 52-46 kya14.
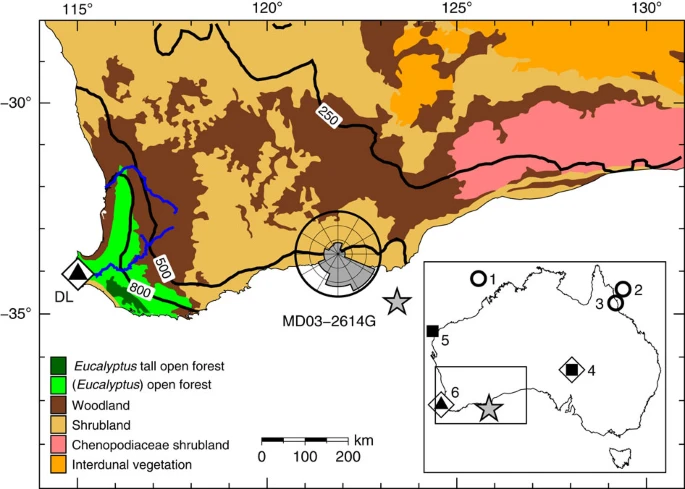
The authors of the Caledonia Fen paper caution that the time scale for their site was not fully resolved (hence the range). Also, it is possible that the timing of fire and vegetation changes at Lynch’s crater took place slightly earlier15 than suggested by Johnson et al. (which I speculate may apply to the timing of spore collapse there too). Regardless, true dates will not be very far off and these studies took care to avoid biased results in dung fungal analysis.
In each case, dung fungi levels fluctuated to some degree but remained high overall, and did not fully recover after crashing for the rest of the Pleistocene. Crucially, extinction estimates from fungal analysis are not far from those derived from reliable fossil analysis suggesting that extinctions were mostly complete by ~41 kya16. There has been difficulty in precisely dating some fossils and sediments due to their considerable ages, but an extinction pulse between 50 and 40 kya is near-certain. This shows that megafaunal collapse in Australia closely followed human entry and was abrupt rather than staggered, even if final extinctions of some species possibly occurred later (though this too would need to be demonstrated).
Glaring Problems in Climatic Arguments
Now that we have taken the crucial step of narrowing down the timeframe for extinctions, we can test the validity of the climate hypothesis. Aridification, or increasing dryness over time, is the form of climate change cited to explain Australia’s extinctions. It serves as a convenient climatic explanation for two reasons17. First, Australia is an already dry continent and it is theoretically plausible that a substantial increase in aridity -gradual or sudden- could push its animals over a hypothetical edge, mainly by diminishing water supplies. Second, the extinctions occurred during a glacial period, and a common but outdated belief is that glacials were markedly drier than interglacials almost everywhere.
A commonly cited study by Hocknull et al. argues that “sustained environmental deterioration” due to aridification from ~48 kya to ~30 kya was the cause of megafaunal collapse18. The authors cast doubt on the reliability of dung fungal analysis, and date sediments associated with megafauna to claim that extinctions occurred after 40 kya. Yet, in the previous section we pointed out that dung fungal analysis can be reliable and indicated megafaunal collapse by 40 kya, not after. Something must have happened just before 40 kya that caused dung fungi concentrations to suddenly fall and not recover whereas they remained high through previous periods. So what can we say about climate change in Australia during the actual extinction window from 50-40 kya?
We must first acknowledge that understanding scale is crucial: demonstrating evidence of some climate change does not prove it was extreme enough to cause widespread megafaunal loss. Given a timeframe of many thousands of years and a landmass as large as Australia, one will inevitably find some evidence of localized climate change. This occurs even in our stable Holocene. Those who advance the aridity hypothesis, therefore, cannot simply cherry-pick examples of regional drying trends but must prove that aridification was both severe and ubiquitous enough to cause sudden continent-wide extinctions of several genera, many of which were dry-adapted19.
No evidence exists for such a drastic climate shift; paleoclimate simulations do not indicate anomalous aridity in Australia from 50-40 kya20. A hydroclimate analysis (involving water bodies) suggests slightly wetter conditions overall than present21. While it is true that the monsoonal region of north Australia experienced a drying trend ~48 kya, it remained wetter than it is at present for thousands of years afterward. Currently, the region is wet enough to support crocodilians. Overall continental drying only set in after 40 kya (following dung fungi decline), and there is no proof that it was unusually severe.
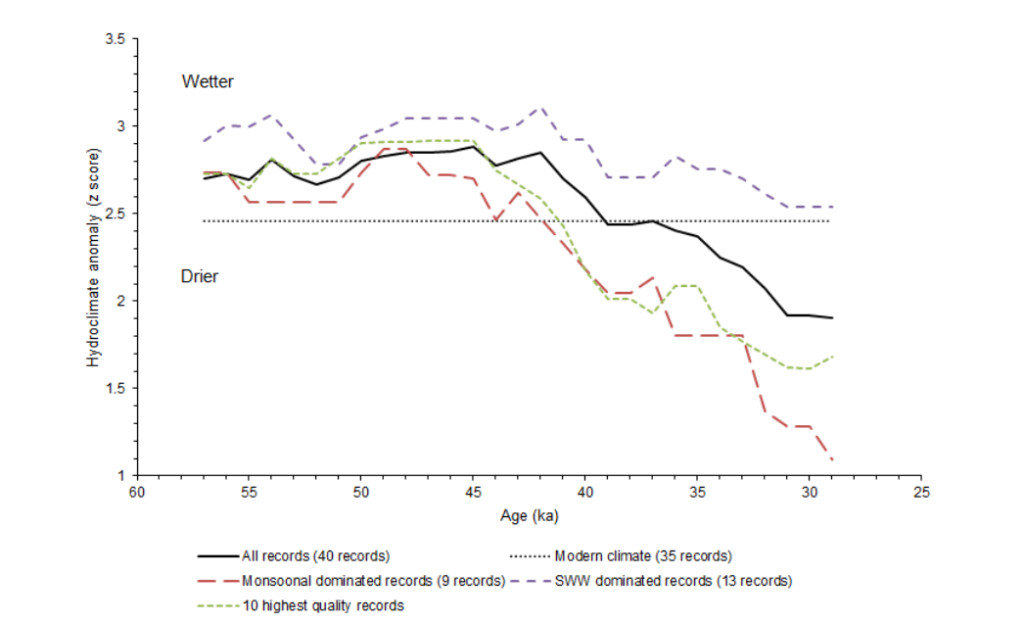
Recent research combining speleothem data with insights into moisture balance and CO2-plant dynamics shows that glacial periods in Australia were not invariably drier than interglacials but alternated between wet and dry phases, while interglacials were consistently dry22. Lower CO2 rather than higher aridity explains reduced woody cover in Australia during glacials. The implication is important: while there being fewer woody plants may have affected some browsing animals, cool conditions during the extinction window did not automatically mean reduced water sources, as confirmed by hydrological studies20.
Further, we would do well to acknowledge that climate patterns on our planet are interconnected-extreme climate change on one continent will happen in the context of major global or hemispheric changes, and will tend to be associated with major shifts in temperature and/or atmospheric circulation patterns. It is therefore unlikely that one continent would experience sudden hyper-aridification while the rest of the Southern Hemisphere remained fairly stable. Studies such as the one by Hocknull et al.18 fail to explain how the purported severe climatic downturn in Australia (48–30 kya) was unprecedented in the Late Pleistocene or aligns with global or hemispheric trends, which is necessary considering the strong evidence pointing to the earlier half of this period being decisive for megafauna.
Yet, perhaps the most overlooked aspect of the climate debate is what we can glean from human history. We must keep in mind that humans were not passive bystanders to acute environmental change. They were affected just as animals were. This is especially the case when it comes to increased aridity, as humans are obviously highly reliant on water. If there was a surge in dryness coinciding with or post-dating human arrival that decimated most Australian megafauna, we would expect to see evidence that resident humans were negatively impacted too.
We do not see this. In fact, genetic testing indicates that the effective population of Aboriginal Australians remained remarkably stable and actually slowly increased after arrival, with no bottleneck anywhere in the past 50,000 years3 23. We also see strong regional differentiation which indicates that Aboriginal groups largely descend from the same ancestors who migrated to their respective regions very long ago, in stark contrast to hunter-gatherers in Europe who faced a combination of extinctions, range contractions, bottlenecks, and population turnovers associated with climatic turbulence24.
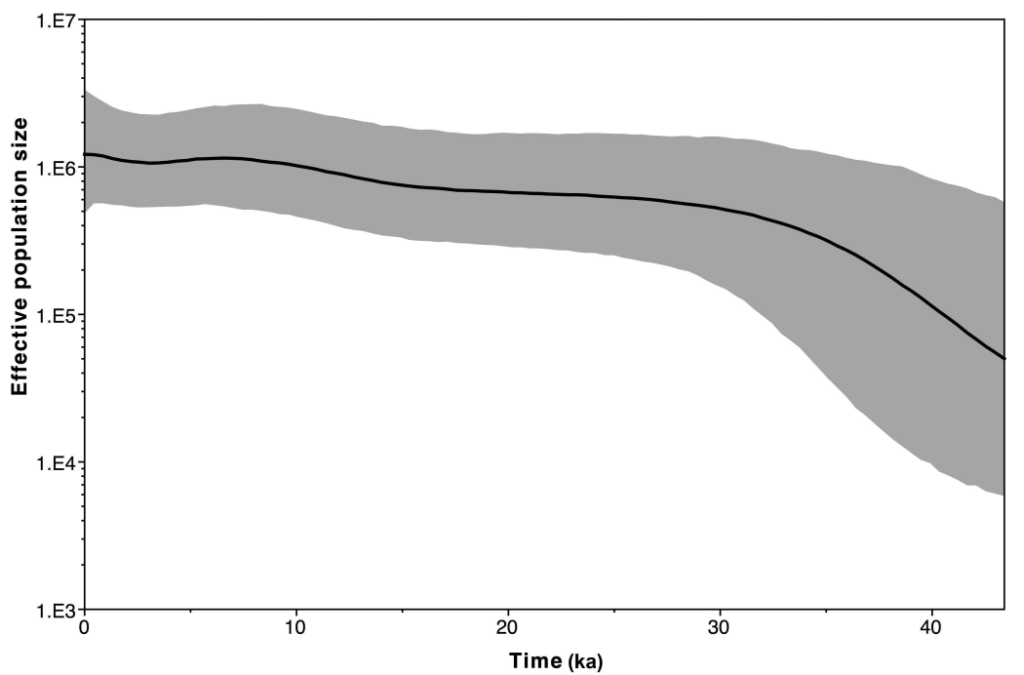
All animals have their limits and even those Australian megafauna whose ancestors had evolved to endure aridity over millions of years were not immune to extreme hyper-aridity. However, attributing their demise to drying raises the question: why did the same aridity fail to eliminate or even affect the newly arrived humans? People were obviously not as physically adapted to surviving on minimal water intake as the animals were, and they must have been getting their water from the same places that animals got it from, as wells were not yet invented.
The more plausible explanation is that Australia was unlikely to have become so arid during the Late Pleistocene that the early human settlers, and by extension the megafauna, were unable to find drinking water or food. The implication is that early Australians simply faced low environmental turbulence, particularly compared to their Pleistocene European counterparts, which fits with Australia being a place of relative climatic stability for the past few million years 25 (see graph below).

Moreover, while every continent, including Australia, experiences natural environmental change and extinction over time, this relative stability casts serious doubt on the hypothesis set forth by Wroe and others wherein heavy, staggered losses of Australian megafauna took place from the Middle Pleistocene onwards 6 18. Such high attrition of megafauna across the same period is not known from Europe and North America where climatic instability was far greater. It would imply the unlikely scenario that Australia followed a uniquely intense pattern of decline despite being located in the climatically more benign Southern Hemisphere and having no major glaciations.
So far in this piece, arguments for climate change as the culprit in extinctions seem quite weak, but there are still more flaws which should be noted. For example, a vast continental shelf was exposed in northern Australia ~50 kya, which continued to expand as sea levels fell further. A critical review by Brook et al. points out that these animals should have been able to shift their ranges away from the arid interior and onto the less-dry continental shelf in the event of severe aridity 26. This seems like another rather obvious issue with climatic arguments (in fact, it had occurred to me independently prior to reading the review).
In fact, we can demonstrate that this shelf was likely a valuable refuge for megafauna by turning yet again to human population genetics. Unlike mainland Australian Aboriginals, the demographics of Tiwi and Elcho islanders off the coast of northern Australia were not stable through prehistory. They reached peak effective population sizes during the Last Glacial Maximum when the land area of the north Australian shelf was greatest and then saw large declines as sea levels rose27. The fact that people thrived when the shelf was most exposed indicates that the land was perfectly suitable for life, which raises the question of why megafauna could not have utilized it if needed. It is not logical that animals would make themselves sitting ducks to climate change by not exploiting the extra real estate available at the time.
Lastly, if aridity was to blame for the extinctions, why were Tasmania and New Guinea not spared the loss of megafauna? Tasmania and especially New Guinea are much wetter than Australia on average. Rain-drenched New Guinea continuously hosted dense rainforests and many rivers28. Yet, its megafauna suffered the same fate as their mainland Australian counterparts following human arrival26, although there is a possibility that they survived significantly longer than their Australian counterparts owing to the sheer remoteness and extraordinarily low human density in New Guinea in prehistoric times 29. Meanwhile, Tasmania’s megafauna immediately following the formation of a land bridge that allowed humans to cross 30. To my knowledge, this inconvenient fact has never been addressed.

The various lines of evidence against intense, widespread drying on mainland Australia, the presence of continental shelves which could mitigate the worst effects of environmental change, and similar extinctions on nearby wet regions indicate climate played at best a minor role in the disappearance of Australian megafauna, likely restricted to a handful of locations. But before wrapping this section up, I will add a final thought-provoking point as a cherry on top.
The red kangaroo (Osphranter rufus) is currently the largest land animal in all of Australasia with males weighing around 70 kg on average. Climate theories would imply that this state of affairs is natural, as all terrestrial megafauna larger than this species succumbed to climate change in the Pleistocene. Yet, there are much larger animals than red kangaroos living in actual deserts in Africa and Asia (donkeys, camels, ostriches, oryx), and in islands like Madagascar prior to human contact (giant lemurs and elephant birds).
In other words, medium-large and large megafauna were able to persist into modern times in resource-limited environments like deserts and islands. Given that context, those espousing climatic causes for the extinction of Australian megafauna must seriously reflect on whether or not the bizarrely diminutive size of Holocene land animals in Australasia-with its many habitats and climate zones-is truly the result of natural processes or not.
It is worth noting that many of the points raised above apply equally well against a long (Middle Pleistocene through Late Pleistocene) or short (between 50-40 kya) chronology for extinction on Sahul.
- Continental shelf exposure: the shelf was exposed during all past glacial intervals, so if glacial aridity did indeed affect in the interior, it could well have compensated for it
- Megafaunal vs human survival: if even dry-adapted Australian megafauna were going extinct due to intense waves of aridity during the mid-to-late Pleistocene, then either an extinction or bottleneck should have affected Aboriginals, as they would have experienced the full range of glacial-interglacial climatic variation during the 50 thousand or so years that they have lived on the continent
- New Guinea and Tasmania: these places have remained either very wet or at least never been particularly dry at any point during the Pleistocene, so the failure of animals to survive here as well indicates something else was the cause
- Sahul as an exception: all other continents and large islands supported diverse megafauna, including numerous large terrestrial species, until the arrival of humans. Even arid regions hosted sizeable animals. It is difficult to explain through natural causes why Sahul, despite its wide range of environments, ended up with such a depauperate and small-bodied terrestrial megafauna. This is especially strange given its location in the Southern Hemisphere and lack of extensive glaciation, which would cause climate changes to be relatively milder
All of these issues make climatic arguments extraordinarily weak. This leaves us with humans as the main culprit as they were the only other agent in the picture that could be responsible for the extinctions. We will explore the anthropogenic dimension next.
Sahulian Susceptibility
Obviously Sahul’s megafauna, like those in the Americas and on islands, had not encountered humans before which was a major factor in their vulnerability. But Sahul was settled quite early on following the human expansion out of Africa, before many of the technological advances of the Upper Paleolithic took place. The hunter gatherers of Sahul had a low population density and their hunting weapons were initially primitive, likely with wooden rather than stone-tipped points 31. So how did they wipe out all those megafauna? I would argue other circumstances specific to Sahul were decisive.
Recall the high degree of isolation of Australasia and dominance of marsupials there. Isolation reduces selection pressure and may hinder competitiveness. Meanwhile, marsupial anatomy itself may preclude marsupial predators from being effective 32, and also results in a lack of cursoriality (running ability) among marsupials generally 33. This could imply that large Sahulian herbivores lacked strong anti-predatory defenses, such as being able to escape quickly, compared to their Afro-Eurasian counterparts. Studies on Procoptodon goliah locomotion seem to validate this 33.
This would not bode well for the megafauna who were about to encounter a super-predator in the form of Homo sapiens. The people who reached Sahul initially lacked the advanced stone tools of their counterparts in Europe and northern Asia 2, but this was offset (perhaps considerably so) by the fact that the prey they encountered were unusually vulnerable. Marsupial and reptilian predators, accustomed to this less challenging milieu, would have been easily outcompeted.

Models suggest a low-density Aboriginal population could quickly drive large mammals like Diprotodon to extinction 31. Nearby New Zealand provides a concrete example of tiny human populations causing the resident megafauna-the Moa-to perish rapidly 35. Lastly, humans would not have been randomly distributed across a landmass as dry as Australia but rather concentrated near water bodies, where they were most likely to cross paths with megafauna 36.
All of this combines to explain why the early settlers were capable of eliminating megafauna through hunting, but this piece would not be complete without mentioning the potentially significant role of Aboriginal fire-use in the extinctions. I consider this one of the most fascinating parts of the extinction debate.
Scorched Earth?
Around the time of human settlement, vegetation change occurred in some places-sclerophyllous (tough leaved) trees like Eucalyptus replaced non-sclerophyllous ones, while the proportion of herbaceous taxa increased. While some view it as proof of climate change, many scientists hypothesize that the shift in vegetation was instead brought on by an increase in fire due to humans. Anthropogenic firing may have replaced fire-sensitive plants with fire-tolerant ones and even partly contributed to the extinctions 37.
Indigenous Australians practiced what is known as fire-stick farming for thousands of years. In The Biggest Estate on Earth: How Aborigines Made Australia, Bill Gammage describes how the Australian landscape looked neat and park-like to early Europeans due to firestick farming, revealing how effectively a low-density human population could wield fire. It was used for clearing vegetation through burning to facilitate hunting by promoting open environments, as well as reducing fire intensity in natural fires by removing excess vegetation.

The extent of anthropogenic fire during the initial human settlement of Sahul, their impacts on its landscapes, and their role in the extinctions remains hotly contested. Some claim that peaks in fire activity after human arrival were natural 38 or that human fire had modest impacts on biota 39. While bushfires have always taken place in Sahul, not all fire may have been natural. Burning and canopy opening occurred even in the wet New Guinea region at times of cool temperatures and relatively high precipitation when bushfires would have been quite rare, possibly indicating human agency 40.
A key site for studying fire and vegetation change is Lynch’s crater in northeast Australia. Initial studies suggested anthropogenic fire preceded the shift from mixed rainforest/sclerophyllous forest to homogenous sclerophyll forest 15. However, later research argued that vegetation change and fire followed megafaunal elimination by humans, as the absence of herbivores allowed excessive plant growth, intensifying natural fires by providing extra fuel 11 41.
At Murray-Darling, a spike in charcoal levels slightly after 50 kya was likely due to humans. Animals went extinct soon after but vegetation did not change appreciably aside from a temporary decline in plants with large seeds and fruits thought to be dispersed by large herbivores 13. The authors conclude that continued fires after extinction were due not to increased vegetative fuel but ongoing burning by people. The results and conclusion are quite different from the second study on Lynch’s crater, but it may be expected as the Murray-Darling basin contained dry, fire tolerant plants as opposed to the wetter, fire-sensitive rainforest taxa of the former.
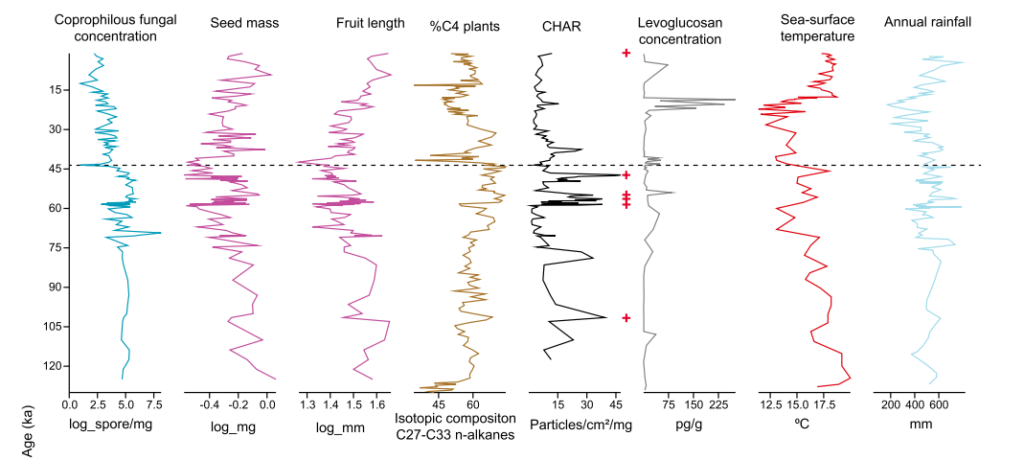
This shows it is necessary to examine ecosystem-specific reactions to fire. The lead author of the Murray-Darling paper produced another great study which did just that by comparing two sites in Tasmania: one located in what was then dry shrubland and another in what was then rainforest 42. It found that the shrubland site saw no vegetative turnover at the outset of human burning around 41 kya, but the rainforest was transformed into a scrubby landscape. The role of herbivore collapse on vegetation was not explored, but human firing probably had the greater impact on flora as burning clearly preceded turnover of rainforest taxa.
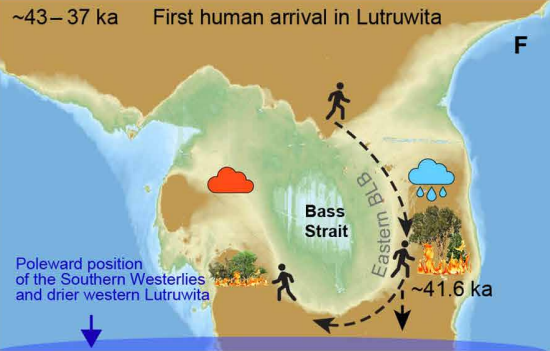
showing more intense fires in the eastern rainforest than in the western scrub
If rainforests(and possibly other wet habitats) were destroyed by humans through burning, this could provide a useful mechanism for extinction. One may speculate that extinction via habitat destruction rather than direct hunting alone could explain why there are no “kill sites” showing evidence of humans hunting megafauna in Australia 6. However, the sheer time gap to the present could be a sufficient explanation, as this makes the chances of such evidence being preserved astronomically low 43.
Further, it is worth noting that most Australian megafauna lived in fire-tolerant habitats but extinctions occurred there too, such as in the Murray-Darling basin 13. Meanwhile, in other areas like Caledonia Fen, no notable increase in fire was recorded around the time of extinction (possibly due to cold conditions of the locale) 14. Moreover, New Guinea’s rainforest persisted through the end of the Pleistocene but its megafauna did not. This could imply hunting was a bigger factor than burning in extinction. More research from more areas, along with more precise chronology will help resolve the exact role of anthropogenic fire in Sahul.
Conclusion
It is not hard to demonstrate why climatic explanations for Late Pleistocene extinctions in Australia fall short, especially focusing on the lack of signs of aridification during the extinctions. It can even be argued that there is irony in searching for evidence of increased dryness as the cause, since any aridification severe enough to wipe out so many types of dry-adapted megafauna across the entirety of Australia would already be known as it would have left clear and unmistakable evidence.
Humans, meanwhile, were more than capable of decimating the vulnerable dwellers of this isolated land despite primitive technology and low population density. They swept across Australia from the north in rapid fashion shortly after 50 kya, with extinction following soon after. The extent to which anthropogenic fire affected flora and contributed to extinctions is uncertain as of yet.
The case of Australia is central to the topic of Late Pleistocene extinctions, and can provide useful insights in the broader debate. Natural environmental change undoubtedly contributed to extinction in large areas of the planet, but Australia and Sahul in general may present the clearest example of prehistoric humans inflicting continent-wide losses of megafauna without much aid from climate change. Recognizing human potency and accounting for continent-specific circumstances will be key to properly assessing the Late Pleistocene extinctions globally.
References
1. Roberts, R. G., Flannery, T. F., Ayliffe, L. K., Yoshida, H., Olley, J. M., Prideaux, G. J., Laslett, G. M., Baynes, A., Smith, M. A., Jones, R., & Smith, B. L. (2001). New Ages for the Last Australian Megafauna: Continent-Wide Extinction About 46,000 Years Ago. Science, 292(5523), 1888–1892. https://doi.org/10.1126/science.1060264
2. Reich, D. (2019). Who we are and how we got here : ancient DNA and the new science of the human past. Oxford University Press.
3. Tobler, R., Rohrlach, A., Soubrier, J., Bover, P., Llamas, B., Tuke, J., Bean, N., Abdullah-Highfold, A., Agius, S., O’Donoghue, A., O’Loughlin, I., Sutton, P., Zilio, F., Walshe, K., Williams, A. N., Turney, C. S. M., Williams, M., Richards, S. M., Mitchell, R. J., & Kowal, E. (2017). Aboriginal mitogenomes reveal 50,000 years of regionalism in Australia. Nature, 544(7649), 180–184. https://doi.org/10.1038/nature21416
4. Allen, J., & O’Connell, J. (2014). Both half right:Updating the evidence for dating first human arrivals in Sahul. Australian Archaeology, 79(1), 86–108. https://doi.org/10.1080/03122417.2014.11682025
5. O’Connell, J. F., Allen, J., Williams, M. A. J., Williams, A. N., Turney, C. S. M., Spooner, N. A., Kamminga, J., Brown, G., & Cooper, A. (2018). When did Homo sapiens first reach Southeast Asia and Sahul? Proceedings of the National Academy of Sciences, 115(34), 8482–8490. https://doi.org/10.1073/pnas.1808385115
6. Wroe, S., Field, J. H., Archer, M., Grayson, D. K., Price, G. J., Louys, J., Faith, J. T., Webb, G. E., Davidson, I., & Mooney, S. D. (2013). Climate change frames debate over the extinction of megafauna in Sahul (Pleistocene Australia-New Guinea). Proceedings of the National Academy of Sciences, 110(22), 8777–8781. https://doi.org/10.1073/pnas.1302698110
7. Trueman, C. N. G., Field, J. H., Dortch, J., Charles, B., & Wroe, S. (2005). Prolonged coexistence of humans and megafauna in Pleistocene Australia. Proceedings of the National Academy of Sciences, 102(23), 8381–8385. https://doi.org/10.1073/pnas.0408975102
8. Brook, B. W., Bradshaw, C. J. A., Cooper, A., Johnson, C. N., Worthy, T. H., Bird, M., Gillespie, R., & Roberts, R. G. (2013). Lack of chronological support for stepwise prehuman extinctions of Australian megafauna. Proceedings of the National Academy of Sciences, 110(36), E3368–E3368. https://doi.org/10.1073/pnas.1309226110
9. David, B., Arnold, L. J., Delannoy, J.-J., Fresløv, J., Urwin, C., Petchey, F., McDowell, M. C., Mullett, R., Mialanes, J., Wood, R., Crouch, J., Berthet, J., Wong, V. N. L., Green, H., & Hellstrom, J. (2021). Late survival of megafauna refuted for Cloggs Cave, SE Australia: Implications for the Australian Late Pleistocene megafauna extinction debate. Quaternary Science Reviews, 253, 106781. https://doi.org/10.1016/j.quascirev.2020.106781
10. Pym, F. C., Franco-Gaviria, F., Espinoza, I. G., & Urrego, D. H. (2023). The timing and ecological consequences of Pleistocene megafaunal decline in the eastern Andes of Colombia. Quaternary Research, 114, 1–17. https://doi.org/10.1017/qua.2022.66
11. Johnson, C. N., Rule, S., Haberle, S. G., Turney, C. S. M., Kershaw, A. P., & Brook, B. W. (2015). Using dung fungi to interpret decline and extinction of megaherbivores: problems and solutions. Quaternary Science Reviews, 110, 107–113. https://doi.org/10.1016/j.quascirev.2014.12.011
12. Van der Kaars, S., Miller, G. H., Turney, C. S. M., Cook, E. J., Nürnberg, D., Schönfeld, J., Kershaw, A. P., & Lehman, S. J. (2017). Humans rather than climate the primary cause of Pleistocene megafaunal extinction in Australia. Nature Communications, 8(1). https://doi.org/10.1038/ncomms14142
13. Adeleye, M. A., Charles Andrew, S., Gallagher, R., van der Kaars, S., De Deckker, P., Hua, Q., & Haberle, S. G. (2023). On the timing of megafaunal extinction and associated floristic consequences in Australia through the lens of functional palaeoecology. Quaternary Science Reviews, 316, 108263. https://doi.org/10.1016/j.quascirev.2023.108263
14. Johnson, C. N., Rule, S., Haberle, S. G., Kershaw, A. P., McKenzie, G. M., & Brook, B. W. (2015). Geographic variation in the ecological effects of extinction of Australia’s Pleistocene megafauna. Ecography, 39(2), 109–116. https://doi.org/10.1111/ecog.01612
15. Turney, C. S. M., Kershaw, A. P., Moss, P., Bird, M. I., Fifield, L. K., Cresswell, R. G., Santos, G. M., Di Tada, M. L., Hausladen, P. A., & Zhou, Y. (2001). Redating the onset of burning at Lynch’s Crater (North Queensland): implications for human settlement in Australia. Journal of Quaternary Science, 16(8), 767–771. https://doi.org/10.1002/jqs.643
16. Gillespie, R., Brook, B. W., & Baynes, A. (2006). Short overlap of humans and megafauna in Pleistocene Australia. Alcheringa: An Australasian Journal of Palaeontology, 30(sup1), 163–186. https://doi.org/10.1080/03115510609506861
17. Field, J., & Wroe, S. (2012). Aridity, faunal adaptations and Australian Late Pleistocene extinctions. World Archaeology, 44(1), 56–74. https://doi.org/10.1080/00438243.2012.647572
18. Hocknull, S. A., Lewis, R., Arnold, L. J., Pietsch, T., Joannes-Boyau, R., Price, G. J., Moss, P., Wood, R., Dosseto, A., Louys, J., Olley, J., & Lawrence, R. A. (2020). Extinction of eastern Sahul megafauna coincides with sustained environmental deterioration. Nature Communications, 11(1). https://doi.org/10.1038/s41467-020-15785-w
19. Prideaux, G. J., Long, J. A., Ayliffe, L. K., Hellstrom, J. C., Pillans, B., Boles, W. E., Hutchinson, M. N., Roberts, R. G., Cupper, M. L., Arnold, L. J., Devine, P. D., & Warburton, N. M. (2007). An arid-adapted middle Pleistocene vertebrate fauna from south-central Australia. Nature, 445(7126), 422–425. https://doi.org/10.1038/nature05471
20. Saltré, F., Rodríguez-Rey, M., Brook, B. W., Johnson, C. N., Turney, C. S. M., Alroy, J., Cooper, A., Beeton, N., Bird, M. I., Fordham, D. A., Gillespie, R., Herrando-Pérez, S., Jacobs, Z., Miller, G. H., Nogués-Bravo, D., Prideaux, G. J., Roberts, R. G., & Bradshaw, C. J. A. (2016). Climate change not to blame for late Quaternary megafauna extinctions in Australia. Nature Communications, 7(1). https://doi.org/10.1038/ncomms10511
21. Kemp, C. W., Tibby, J., Arnold, L. J., & Barr, C. (2019). Australian hydroclimate during Marine Isotope Stage 3: A synthesis and review. Quaternary Science Reviews, 204, 94–104. https://doi.org/10.1016/j.quascirev.2018.11.016
22. Weij, R., Sniderman, J. M. K., Woodhead, J. D., Hellstrom, J. C., Brown, J. R., Drysdale, R. N., Reed, E., Bourne, S., & Gordon, J. (2024). Elevated Southern Hemisphere moisture availability during glacial periods. Nature, 626(7998), 319–326. https://doi.org/10.1038/s41586-023-06989-3
23. Pedro, N., Brucato, N., Fernandes, V., André, M., Saag, L., Pomat, W., Besse, C., Boland, A., Deleuze, J.-F., Clarkson, C., Sudoyo, H., Metspalu, M., Stoneking, M., Cox, M. P., Leavesley, M., Pereira, L., & Ricaut, F.-X. (2020). Papuan mitochondrial genomes and the settlement of Sahul. Journal of Human Genetics, 65(10), 875–887. https://doi.org/10.1038/s10038-020-0781-3
24. Posth, C., Yu, H., Ghalichi, A., Rougier, H., Crevecoeur, I., Huang, Y., Ringbauer, H., Rohrlach, A. B., Nägele, K., Villalba-Mouco, V., Radzeviciute, R., Ferraz, T., Stoessel, A., Tukhbatova, R., Drucker, D. G., Lari, M., Modi, A., Vai, S., Saupe, T., & Scheib, C. L. (2023). Palaeogenomics of Upper Palaeolithic to Neolithic European hunter-gatherers. Nature, 615(7950), 117–126. https://doi.org/10.1038/s41586-023-05726-0
25. Herrando-Moraira, S., Nualart, N., Galbany-Casals, M., Garcia-Jacas, N., Ohashi, H., Matsui, T., Susanna, A., Tang, C. Q., & López-Pujol, J. (2022). Climate Stability Index maps, a global high resolution cartography of climate stability from Pliocene to 2100. Scientific Data, 9(1), 48. https://doi.org/10.1038/s41597-022-01144-5
26. Brook, B. W., Bowman, D. M. J. S., Burney, D. A., Flannery, T. F., Gagan, M. K., Gillespie, R., Johnson, C. N., Kershaw, P., Magee, J. W., Martin, P. S., Miller, G. H., Peiser, B., & Roberts, R. G. (2007). Would the Australian megafauna have become extinct if humans had never colonised the continent? Comments on “A review of the evidence for a human role in the extinction of Australian megafauna and an alternative explanation” by S. Wroe and J. Field. Quaternary Science Reviews, 26(3), 560–564. https://doi.org/10.1016/j.quascirev.2006.10.008
27. Silcocks, M., Farlow, A., Hermes, A., Tsambos, G., Patel, H. R., Huebner, S., Baynam, G., Jenkins, M. R., Vukcevic, D., Easteal, S., & Leslie, S. (2023). Indigenous Australian genomes show deep structure and rich novel variation. Nature, 624(7992), 593–601. https://doi.org/10.1038/s41586-023-06831-w
28. Summerhayes, G. R., Field, J. H., Shaw, B., & Gaffney, D. (2017). The archaeology of forest exploitation and change in the tropics during the Pleistocene: The case of Northern Sahul (Pleistocene New Guinea). Quaternary International, 448, 14–30. https://doi.org/10.1016/j.quaint.2016.04.023
29. Prideaux, G. J., Kerr, I. A. R., van Zoelen, J. D., Grün, R., van der Kaars, S., Oertle, A., Douka, K., Grono, E., Barron, A., Mountain, M., Westaway, M. C., & Denham, T. (2022). Re‐evaluating the evidence for late‐surviving megafauna at Nombe rockshelter in the New Guinea highlands. Archaeology in Oceania, 57(3), 223–248. https://doi.org/10.1002/arco.5274
30. Turney, C. S. M., Flannery, T. F., Roberts, R. G., Reid, C., Fifield, L. K., Higham, T. F. G., Jacobs, Z., Kemp, N., Colhoun, E. A., Kalin, R. M., & Ogle, N. (2008). Late-surviving megafauna in Tasmania, Australia, implicate human involvement in their extinction. Proceedings of the National Academy of Sciences, 105(34), 12150–12153. https://doi.org/10.1073/pnas.0801360105
31. Brook, B., & Johnson, C. (2006). Selective hunting of juveniles as a cause of the imperceptible overkill of the Australian Pleistocene megafauna. Alcheringa: An Australasian Journal of Palaeontology, 31, 39–48. https://doi.org/10.1080/03115510608619573
32. Faurby, S., & Svenning, J.-C. (2016). The asymmetry in the Great American Biotic Interchange in mammals is consistent with differential susceptibility to mammalian predation. Global Ecology and Biogeography, 25(12), 1443–1453. https://doi.org/10.1111/geb.12504
33. Martin, M. L., & Weisbecker, V. (2023). Function and Constraint in the Marsupial Postcranium. Springer EBooks, 1–27. https://doi.org/10.1007/978-3-030-88800-8_49-1
34. McGowan, C. P., Skinner, J., & Biewener, A. A. (2008). Hind limb scaling of kangaroos and wallabies (superfamily Macropodoidea): implications for hopping performance, safety factor and elastic savings. Journal of Anatomy, 212(2), 153–163. https://doi.org/10.1111/j.1469-7580.2007.00841.x
35. Holdaway, R. N., Allentoft, M. E., Jacomb, C., Oskam, C. L., Beavan, N. R., & Bunce, M. (2014). An extremely low-density human population exterminated New Zealand moa. Nature Communications, 5(1). https://doi.org/10.1038/ncomms6436
36. O’Connell, J.F., 2000. An emu hunt. In: Anderson, A., Murray, T. (Eds.), AustralianArchaeologist: Collected Papers in Honour of Jim Allen. Division of Archaeologyand Natural History, Research School of Pacific and Asian Studies. AustralianNational University, Canberra, pp. 172–181
37. Miller, G. H. (2005). Ecosystem Collapse in Pleistocene Australia and a Human Role in Megafaunal Extinction. Science, 309(5732), 287–290. https://doi.org/10.1126/science.1111288
38. Mooney, S. D., Harrison, S. P., Bartlein, P. J., Daniau, A.-L. ., Stevenson, J., Brownlie, K. C., Buckman, S., Cupper, M., Luly, J., Black, M., Colhoun, E., D’Costa, D., Dodson, J., Haberle, S., Hope, G. S., Kershaw, P., Kenyon, C., McKenzie, M., & Williams, N. (2011). Late Quaternary fire regimes of Australasia. Quaternary Science Reviews, 30(1), 28–46. https://doi.org/10.1016/j.quascirev.2010.10.010
39. Johnson, C. N. (2016). Fire, people and ecosystem change in Pleistocene Australia. Australian Journal of Botany, 64(8), 643. https://doi.org/10.1071/bt16138
40. Lentfer, C., Pavlides, C., & Specht, J. (2010). Natural and human impacts in a 35 000-year vegetation history in central New Britain, Papua New Guinea. Quaternary Science Reviews, 29(27-28), 3750–3767. https://doi.org/10.1016/j.quascirev.2010.08.009
41. Rule, S., Brook, B. W., Haberle, S. G., Turney, C. S. M., Kershaw, A. P., & Johnson, C. N. (2012). The Aftermath of Megafaunal Extinction: Ecosystem Transformation in Pleistocene Australia. Science, 335(6075), 1483–1486. https://doi.org/10.1126/science.1214261
42. Adeleye, M. A., Hopf, F., Haberle, S. G., Stannard, G. L., Mcwethy, D. B., Harris, S., & David. (2024). Landscape burning facilitated Aboriginal migration into Lutruwita/Tasmania 41,600 years ago. Science Advances, 10(46). https://doi.org/10.1126/sciadv.adp6579
43. Surovell, T. A., & Grund, B. S. (2012). The Associational Critique of Quaternary Overkill and why it is Largely Irrelevant to the Extinction Debate. American Antiquity, 77(4), 672–687. https://doi.org/10.7183/0002-7316.77.4.672