Welcome to part 2 of this series on Late Pleistocene American extinctions. In Part 1, we introduced the debate regarding the Late Pleistocene extinctions in the Americas and thoroughly explained why humans were not simply a factor, but the main factor in the demise of megafauna. In this section, we will explore the evidence for human predation on extinct animals, as well as intricately describe a plausible scenario for their extinctions. We will also discuss the importance of possible human-driven environmental change in the context of this topic.
The first order of business, however, is addressing the disagreement over when humans actually arrived in the Americas.
Dating the Arrival of People
In the Americas, there is alleged evidence for human presence dating back to 20,000+ years before present1 2 3 . As extinctions occurred between 14-10 kya, it may be tempting to wonder why it took so long for humans to eliminate megafauna if they truly arrived so early. I will get into why we should not expect immediate extinctions in the case of the Americas, but for now, I want to focus on what we can say about the alleged early sites and their implications.
There is a lot of excitement over early arrival dates for humans in the New World. However, I have written about this subject in a previous post and argued for why we should be skeptical of these claims. Genetic analysis suggests the ancestors of modern Native Americans arrived south of the ice sheets around 16 kya4. This pre-dates Clovis to be sure, but it is quite different from LGM or pre-LGM habitation. A human presence in mid-latitude North America during the LGM(18-22 kya) would imply humans arrived there earlier than the LGM, because both coastal and inland routes from Beringia were blocked during the LGM5.
Additionally, it is tempting to wonder why settlement dates of 20 kya or earlier, if correct, have not led to discoveries of human DNA or human fossils much older than the remains we currently have. In Eurasia, there are a number of sequenced Homo sapiens dating to 30-50 kya, including Tianyuan man and Bacho Kiro, along with much older archaic human remains. Yet, in the Americas, the oldest Paleo-Indian remains seem to date from only 13 kya or so6.
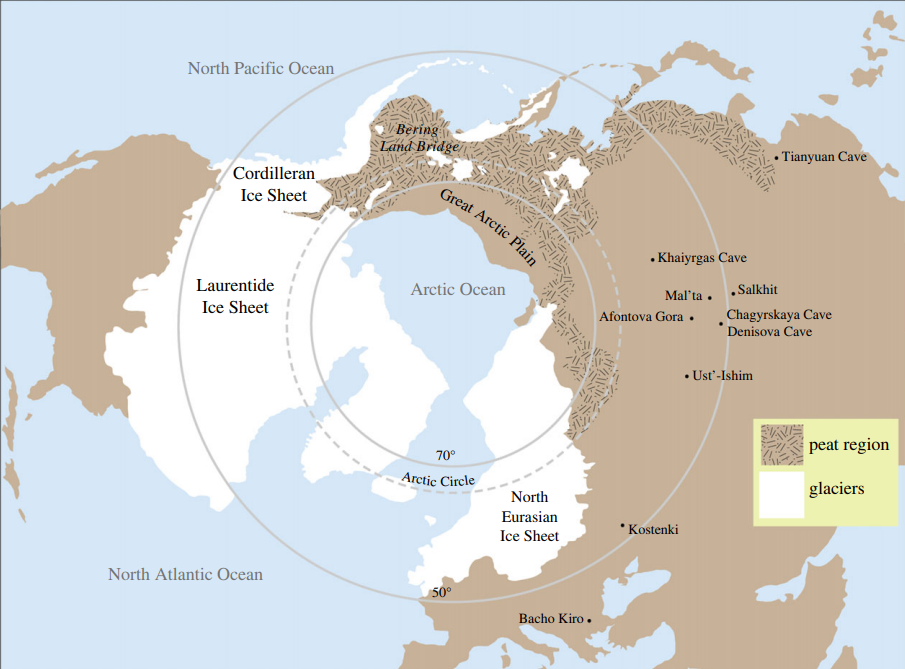
All of this does not categorically refute the idea that there were humans prior to the dates suggested by genomic studies, but it does make it much less likely. If there were, they would not be the direct ancestors of modern Native Americans and instead belong to an unknown population. Also, importantly for the topic of extinction, the lack of human remains from these hypothetical early settlers indicates they must have been far too small in number to appreciably affect wildlife.
With that out of the way, we can discuss what we know about human-megafauna interactions.
Evidence of Human Predation on Megafauna
A major point used by detractors of the overkill hypothesis has been the alleged lack of evidence for hunting of extinct species (especially kill sites)7. It is worth noting that the chances of evidence of hunting being preserved over thousands of years are very small8 9. That said, a number of kill sites are known from across North and South America.
A 2008 study analyzed Clovis-era proboscidean kill sites in North America10. They used the most conservative possible number of 14 sites, all from the United States, based on very stringent standards set by Grayson and Meltzer11. Mammoth and mastodon remains were found next to Clovis projectile points at these sites-an unambiguous sign of interactions. While 14 does not sound like a large number, when the same stringent criteria are used to isolate proboscidean sites all over the Old World where humans were present for much longer, it actually reveals a remarkably strong degree of proboscidean hunting by the Clovis.


In a possible pre-Clovis site in southwestern Canada called Wally’s beach, there is evidence of horse and camel butchery12. 7 horses and 1 camel were killed and consumed near a river at this location which at the time was at the southern edge of the inland ice-free corridor. In the Mexican state of Sonora, there is evidence of Clovis predation on and butchery of a different proboscidean aside from the usual mammoths and mastodons: the gomphothere Cuvieronius 13.


Meanwhile in South America, the Pampas preserve rich evidence of human-megafauna interaction. At Paso Otero 5, the bones of various taxa such as Toxodon, Macrauchenia, Equus, and Glyptodon were found in association with Fishtail projectile points14. The bones were burned, likely for fuel. Another Pampan site, Campo Laborde, represents a confirmed case of human butchery of ground sloth, with cut marks on Megatherium bones indicative of defleshing and skinning15.

Going beyond kill sites, testing of blood residues on Paleo-Indian artifacts in South Carolina shows that the people indeed hunted extinct megafauna16. Proteins in the blood were compared with extant animals to show that they hunted proboscideans and horses. They also hunted ursids (bears) but the paper does not say whether the matches represent extinct or extant members of that genus.
On the other side of the continent in Montana, it was shown via isotopic analysis that the mother of a Clovis child named “Anzick-1” was extremely reliant on megafauna, and mammoth in particular17. 40% of her diet is estimated to have been mammoth, with the rest being mostly elk and (presumably) bison. In fact, isotopes suggest a similar diet to Homotherium, a mammoth specialist. The faunal associations found in the Anzick site are similar to Clovis sites in much of that region, so it can be reasonably inferred that other Clovis in the area had a similar diet.
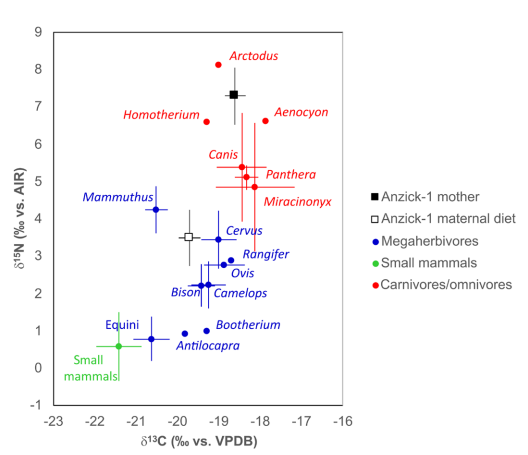
It is important to recognize that the vast majority of dead animal specimens are simply not preserved as fossils9, let alone found, making it very difficult to find evidence of remains that specifically show evidence of predation by humans. This is especially going to be the case if extinction was rapid, as it seems to have been in the Americas9. When considering how rare preservation actually is, there is clearly no shortage of evidence for human hunting of megafauna in North or South America.
Aside from that, we must mention a very interesting and perhaps overlooked implication from one of these findings. If other Paleo-Indians were as or nearly as carnivorous as Anzick-1 and other western Clovis, this could have contributed to or sped up extinction in surprising ways. Anthropologist Miki Ben-Dor has written extensively about how humans being inefficient at digesting protein means that they must rely on lots of fat if pursuing a mostly animal-based diet, leading to leaner, less-choice parts of the animal being wasted18. This wastage implies that humans rely less on any one animal than one might imagine, and would need to kill quite a lot to sustain themselves.
If highly carnivorous Paleo-Indians were forgoing large quantities of lean meat in favor of maximizing the fat-to-protein ratio in their diets, the result might have been intense overhunting. Such overhunting would be in stark contrast to popular notions of hunter-gatherers practicing efficiency and ecological sustainability. Further, it could explain why such a small number of humans had such disproportionate effects on wildlife.
Dispersal, Growth, Collapse
Some researchers speculate on why it may have taken so long for the people to start wiping out megafauna if archeological evidence suggests humans arrived in the Americas south of the ice sheets 16-15 kya19. They consider some possibilities such as the initial settlers being generalists or too low in population to affect animals. I believe these two are the correct answers, but I would like to flesh them out a little more by providing a potential scenario for extinction from beginning to end, and in doing so show why it is reasonable to expect a delay in extinction following arrival.
Please note that there will be heavy speculation involved here, as the manner and timing of settlement in the Americas is not fully resolved as of yet. To summarize the key points in this section:
- Island extinctions are not perfect analogues for extinction events on large continents, and probably give an unreasonable expectation of how fast they should occur.
- The first Americans south of the ice sheets came before Clovis and likely took a Pacific coastal route. They fanned out across the two continents very rapidly.
- Initially, their densities were extremely low, and they may have lacked the numbers, technology, or hunting strategies to bring down megafauna, especially very large species, effectively.
- Rapid population growth aided innovation in hunting technology and possibly hunting strategies, and the combined force led to extinction events. Due to the fact that the settlers had already colonized all regions by this time, megafaunal collapse occurred almost synchronously in South America and mid-latitude North America.
- Clovis projectile points in North America south of the ice sheets and Fishtail points in South America were decisive technological advancements. Meanwhile in eastern Beringia (Alaska/Yukon), where extinctions occurred slightly earlier, microblades were in use.
Starting from the first, the idea that extinction should instantly follow human arrival on a new continent-the blitzkrieg version of overkill- is based on island extinctions as well as the Clovis-first theory20 21. In many islands like New Zealand, humans eliminated the native megafauna immediately after arrival22, which may lead to the assumption that continental extinctions should also be fairly rapid. Meanwhile, the Clovis-first theory posits that the Clovis were the first people in the Americas or at least the first to reach south of the ice sheets. As the Clovis were big-game hunters, it would make sense to assume that they would immediately have an impact on large animals in the Americas.
While island extinctions may provide a useful analogue for what happens when humans encounter unfamiliar landmasses, they are not perfect comparisons when it comes to continents. Island fauna are uniquely vulnerable to extinction in general, and especially extinction at the hands of novel predators. It is not a coincidence that the most “island-like” continent of Australia suffered, proportionally, the worst megafaunal extinctions of any continent following human arrival23. For other continents like North and South America, instant extinction (under 1000 years) should not be considered a given regardless of whether one supports the overkill model or not.
Additionally, the Clovis-first theory and associated land-route hypothesis have been challenged. Both (but especially the former) are increasingly being abandoned by archeologists. Many now believe that the first people south of the ice sheet came somewhat before the Clovis and arrived via a coastal route, after having spent thousands of years in Beringia24 25. Some speculate that while in Beringia, they lived a maritime lifestyle along the shrubby, now-submerged coast rather than the mammoth steppe further north, using the resources of the sea25. A coastal migration by a people adept at exploiting marine resources could explain the early elimination of the Steller’s Sea Cow from the western coast of North America26.
How did the initial Paleo-Indians to make landfall south of the ice spread out? A paper by Gary Haynes speculates about a rapid early-dispersal of the first North Americans, who dispersed from one area to another well before hitting carrying capacity27. In his extinction/settlement model, very small groups of hunter-gatherers spread out across all regions but settled resource-rich areas preferentially, and this initial movement of people hit animal populations hard. Then, successive “shocks” from hunting and climate change finished off the remaining megafauna.
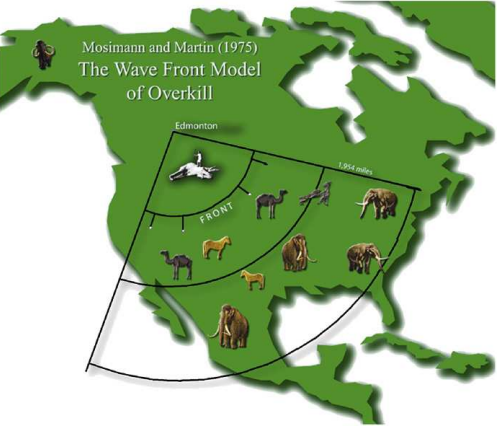
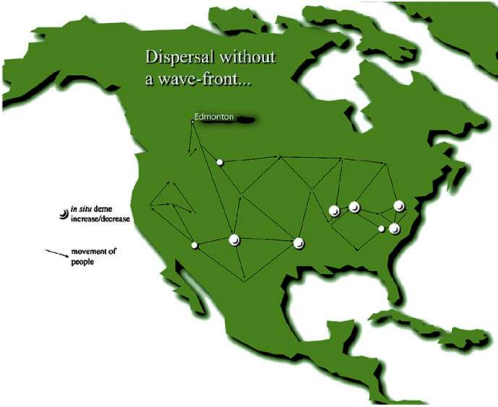
I do not agree with everything in this model, as evidence suggests the initial men to disperse had little effect on megafauna populations28 29, and my last article argues that climate was likely not a necessary factor overall. However, the idea that dispersal occurred in this fashion as opposed to a “wave-front” is something that I believe is very likely. Studies seem to validate that there was indeed a very rapid movement of people across the Americas soon after arrival south of the ice sheets24. Dispersal probably preceded them reaching carrying capacity anywhere, as their population continued to rise afterwards4 29 .
This rapid expansion to the furthest reaches of the New World by approximately 14 kya, and continued growth of the population for thousands of years is nothing short of remarkable. This is in spite of Paleo-Indians encountering far more numerous predators than what their ancestors in southern Beringia did, particularly in temperate North America where the predator guild was notably diverse. In Central and much of South America, the Arctic-origin population had to acclimate to tropical climates and contend with the associated diseases and venomous species. All of this speaks to the incredible adaptability, ingenuity, and fortitude of the ancestors of Native Americans.
In the parts of Beringia that were once covered by mammoth steppe, people were using microblades similar to those from Asia since at least 14.2 kya30. This, along with climate change affecting grazers, could explain why extinctions hit Beringia first31 32. Meanwhile, south of the ice sheets, extinctions started around 13 kya29 31, well after the genetic and archeological evidence for human arrival but coincident with the emergence and spread of Clovis technology from the southern US. This would indicate that the Clovis culture was not the direct heir of Siberian hunters as once thought, but instead represents a people who took up big-game hunting independently.
The authors of one study suggest that Clovis points either evolved from earlier stemmed blades brought by the initial coastal migration or from precursor blades introduced by a second migration from Beringia through the inland ice free corridor24. I lean toward the first scenario, as I would predict that a second migration from Beringia would bring microblades instead of Clovis blade precursors. Regardless, there seems to be an initial hiatus following arrival during which Paleo-Indians were likely generalists without big-game specialist technology.
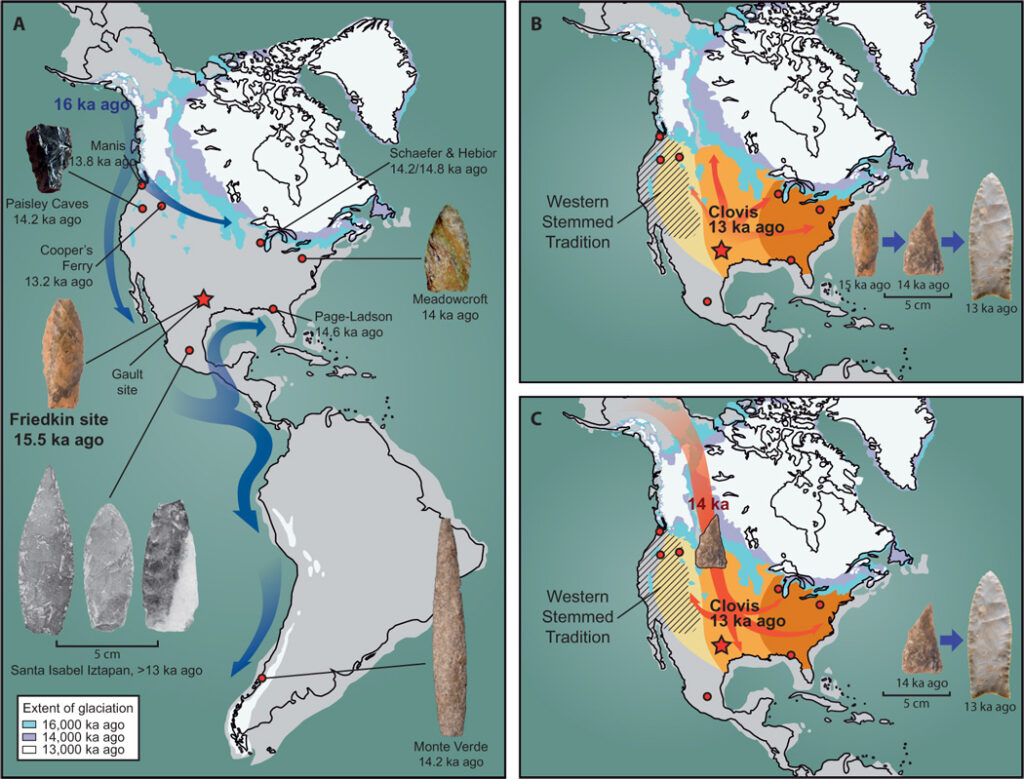
B) One scenario of Clovis point spread from southern US outward
C) Alternate scenario of Clovis pre-cursor spread from Beringia to the south
Waters et al. (2018)
The extinction pulse is strongly linked to the emergence of fluted hunting technology: Clovis and Fishtails in North and South America respectively33. However, in part of western North America, pre-Clovis hunting of horses and camels could have contributed to their early declines12, whereas in the east, horses may have survived into and still been hunted in post-Clovis times16. Also, in the Pampas of South America, non-Fishtail points were still present during the Fishtail era15, indicating that while Clovis and Fishtail points were pivotal developments, technology-extinction dynamics were complex and varied by region.
To better understand what happened in the Americas and why extinction struck when it did, the first 2 minutes and 20 seconds of the clip below featuring world-renowned Harvard geneticist David Reich offers great insights. He describes a feedback loop of population growth and shared knowledge in hunter-gatherer societies, and how eventually a critical mass may be reached which causes runaway progress.
The emergence of continent-wide cultural complexes with potent hunting ability in the Americas like Clovis could be an example of this-something that happened only after previously isolated tribes started exchanging knowledge and technology with each other as population density grew. Human density, knowledge, and technology likely all pushed each other forward slowly until a certain threshold was reached, after which extinction of megafauna likely became an inevitability.
Still, it is important to consider the possibility that Clovis or Fishtail blades were not essential in bringing down megafauna, and that the predecessor and/or successor blade types were just as effective in pursuit of that goal. In that case, it may well be that the actually decisive element here was population growth and/or a cultural shift towards big-game hunting, with the spread of the blade types simply being a signature but non-necessary part of a bigger cultural package. It is difficult to know for sure.
Further, if a major cultural shift did occur, it could have involved the development and spread of new, effective hunting techniques as opposed to new types of weapons alone. A recent article speculated that Clovis people may have used their spears as pikes, held in place to be used against large animals that charged at them 34. If true, this would be an ingenuous method which could partly explain the rapid elimination of proboscideans. Who knows what other innovations in hunting strategy could have taken place as the human population expanded and became better acquainted with their prey?
Anyway, going back to megafaunal collapse: once the populations of herbivores diminished, many of the large predators like sabre-toothed cats, dire wolves, American lions, and short-faced bears that relied on them perished too. The cougar, jaguar, grey wolf, brown bear, and black bear survived, however. How much of the extinction of the large predators was due to direct violent conflict with humans vs. plain loss of food sources is another thing that is hard to determine.
To summarize how extinction may have unfolded in the New World: In Beringia, adverse climate change caused populations of certain animals to decline early on but humans using microblade technology were likely the necessary killing blow for many of them. In the rest of North America as well as South America, Paleo-Indians started using Clovis and Fishtail blades respectively, which marks the point when megafauna populations crashed and eventually perished. The speed and timing of extinction could have been the result of a confluence of population growth, improved technology, and new hunting strategies.
A major difference between Australia on one hand and the Americas on the other is the extreme isolation of the former’s fauna. Therefore, the combined threshold to trigger widespread extinction would be higher in the Americas (especially North America). This explains why the first Australians were able to eliminate megafauna with very primitive weapons. Still, even there, it is possible that there was a slight delay between human arrival and extinction in any given locale, as a result of human density being too low initially. This is difficult to know because a precise chronology for Australia is lacking.
Ultimately, the difference between extinctions on islands and those on continents may just be time. The process of extinction in the Americas should not be assumed to be either linear or immediate, and depended upon multiple factors.
Trophic Networks
As mentioned in the previous part, there are ways in which humans could have triggered extinctions indirectly. One possibility is that they disrupted trophic networks by making animal communities unstable and causing them to collapse.
A 2014 study speculates that Pleistocene North and South American animal communities were vulnerable to perturbations due to how complex they were, and even the simple addition of a novel predator that behaved in a similar fashion to a large predator could be enough to destabilize animal communities and cause them to collapse35. Large predators interact with a wide variety of prey, and if humans behaved similarly, they could have increased connectivity between different species. For example, if people on a new continent eliminated a handful of important prey items, other predators could have start preying heavily on animals they did not normally prey on, causing those animals to go extinct too.
The authors speculate that a larger number of predators would make communities more unstable and a larger number of large herbivores more stable, and speculate that this could be why extinctions took longer in South America, which had less large predators and more large herbivores than North America. Still, extinctions were more severe in South America than North America 23, meaning that while it is possible they influenced the speed of extinction, they had less influence on which animals or what percentage would survive or not. This means the end result is essentially the same.
July 3rd 2026 edit: Contrary to what I initially wrote, extinctions do seem to have taken noticeably longer in South America than North America, even if not to the extent that some have claimed based on erroneously dated specimens.
A Changed Wilderness
In my previous post, I used “environmental changes” to describe shifts in climate and associated changes in vegetation. However, not all environmental changes appear to be driven by climate alone and instead could be linked to the actions of man, which complicates the picture. This may be another potential indirect way that humans drove extinctions in the Americas.
Shifts in fire intensity and vegetation around the time of human arrival and indeed into the Holocene have caused many to speculate about the indirect effects of prehistoric humans on ecosystems. Some argue that megafaunal elimination led to open-canopy ecosystems being replaced by closed-canopy ones, harming smaller species reliant on open habitats36. Others point to increased fire intensity, possibly caused by human-set fires or increased fuel loads after large animals went extinct37 38.
At Rancho La Brea in southern California, a spike in fire around the time of human settlement is speculated to have hastened the transition from open woodland to less-productive chaparral37. In the midwestern and northeastern United States, megafaunal extinction is speculated by some to have contributed to increased fire due to overgrown vegetation 38. In the South American Andes, a similar spike in fire is noted around the time of human arrival 39.
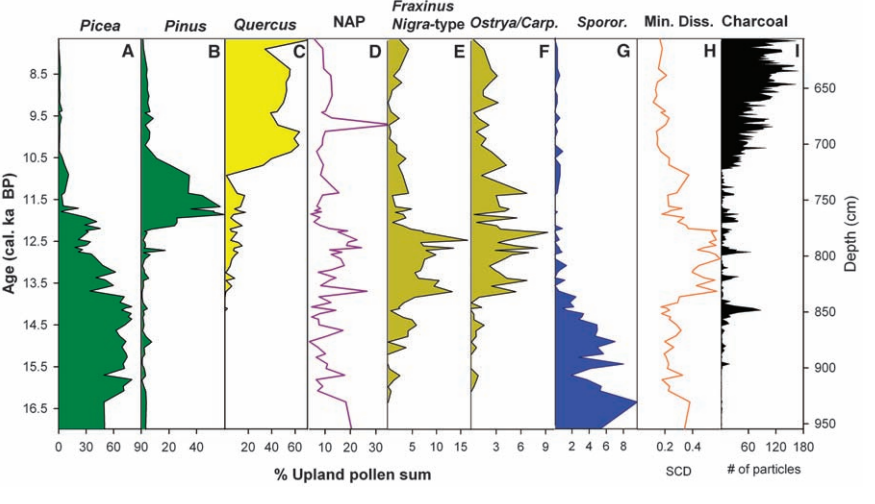
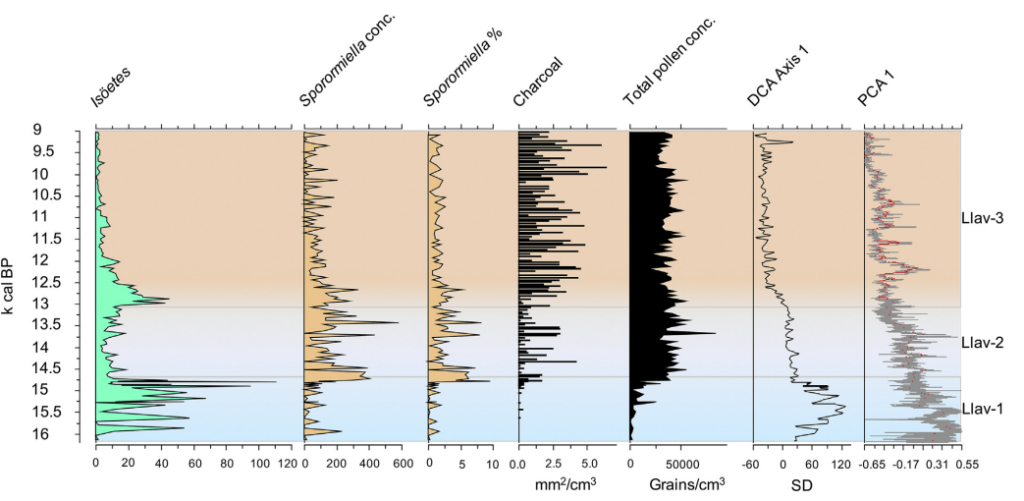
While it may be tempting to attribute this to the effects of climate change alone, especially in southern California where the climate became not only warmer but drier, data comparing past interglacials to the present one in Central and South America indicate that fire levels during the Holocene are unprecedented and had major effects on vegetation40 41. A study comparing past interglacials in the Yukon to the present one also notes the uniqueness of the Holocene, with fire being suggested as the reason for a greater proportion of pine and alder instead of spruce and fir42.
Determining how much of the increased fire during the transition to the Holocene, and the Holocene itself for that matter, resulted from deliberate burning by people vs. heavier vegetative fuel loads after megafaunal extinction remains a challenge. Yet, significant environmental shifts clearly occurred. But did human-driven changes indirectly cause extinctions?
This is a hard question to answer. Going back yet again to Australia, similar speculation exists about the role of fire-use by early settlers in causing extinction 43. However, there is a long history of natural intense fire events in Australia 44, and the majority of Australian ecosystems are arid and fire-resistant. This has led some to question this notion 45. In the Americas meanwhile, multiple fairly humid regions that were not fire prone experienced sudden increase in fire following human arrival 38 39 40, so there is a distinct possibility that unprecedented burning harmed sensitive ecosystems resulting in extinctions.
Certain woody taxa clearly increased in abundance, possibly due to megaherbivore elimination. In one part of California, Alnus, a tree-forming shrub particularly sensitive to animal disturbance, initially declined at the start of the Younger Dryas as expected with sudden cooling but then surged before temperatures warmed46. While the authors of the study do not mention this, it is worth noting that the surge coincided with the end of the Clovis period ~12,700 years ago. Another study shows that Alnus abundance between the Younger Dryas and mid-Holocene was unprecedented in the last 130,000 years47.

some parts of North America after megafaunal loss
Today, closed forests dominate vast regions of the Americas, especially humid zones. Some believe that much of the area occupied by closed forest was once parkland or open woodlands maintained by megafauna, supporting greater biodiversity 36. Recent studies from Europe suggest extinct megafauna maintained habitat mosaics during past interglacials 48 49. It is probable similar dynamics existed in the Americas. The role of canopy closure in driving species to extinction—especially among animals with restricted niches and ranges—cannot be overlooked.
However, specific relationships will need clarification: For example, which species exactly were most vulnerable to increased fire, canopy closure, or other indirect effects? We know that humans did not exclusively hunt large animals, and exploited small megafauna and non-megafauna (like certain birds) too. Could the extinctions of said animals alternatively be explained by direct overhunting (or even climate change for that matter)?
All these questions and more can be answered with more research on vegetation, fire, and animal history spanning long time periods and multiple regions. Even if we cannot establish probable links between habitat change and extinction of certain species, we will at least gain a better understanding of how profoundly what we consider to be “pristine wilderness” has been modified.
Concluding Remarks
The Late Pleistocene extinctions in the Americas, and indeed the rest of the world, are a contentious topic. The debate over whether climate or humans played the larger role will continue for the foreseeable future, but a careful analysis shows that the evidence overwhelmingly points in one direction. We would benefit from more research on how, rather than if, prehistoric Americans strongly impacted animal and plant communities.
The event itself is undoubtedly quite unfortunate but the story of how it happened is immensely fascinating. It should not be used to cast blame or judgment on the earliest Americans, as people living at the time were only trying to survive and probably lacked awareness of how profoundly they were changing ecosystems. Moreover, this is part of a shared history of mankind-humans left deep marks on their environment wherever they went.
It is important to explore this history as accurately as possible and without bias.
References
1. Ardelean, C. F., Becerra-Valdivia, L., Pedersen, M. W., Schwenninger, J.-L., Oviatt, C. G., Macías-Quintero, J. I., Arroyo-Cabrales, J., Sikora, M., Ocampo-Díaz, Y. Z. E., Rubio-Cisneros, I. I., Watling, J. G., de Medeiros, V. B., De Oliveira, P. E., Barba-Pingarón, L., Ortiz-Butrón, A., Blancas-Vázquez, J., Rivera-González, I., Solís-Rosales, C., Rodríguez-Ceja, M., & Gandy, D. A. (2020). Evidence of human occupation in Mexico around the Last Glacial Maximum. Nature, 584(7819), 87–92. https://doi.org/10.1038/s41586-020-2509-0
2. Bennett, M. R., Bustos, D., Pigati, J. S., Springer, K. B., Urban, T. M., Holliday, V. T., Reynolds, S. C., Budka, M., Honke, J. S., Hudson, A. M., Fenerty, B., Connelly, C., Martinez, P. J., Santucci, V. L., & Odess, D. (2021). Evidence of humans in North America during the Last Glacial Maximum. Science, 373(6562), 1528–1531. https://doi.org/10.1126/science.abg7586
3. Thaís Rabito Pansani, Pobiner, B., Guériau, P., Mathieu Thoury, Tafforeau, P., Baranger, E., Águeda Vilhena Vialou, Vialou, D., McSparron, C., Castro, M. C., Trindade, A., Bertrand, L., & Alves, L. (2023). Evidence of artefacts made of giant sloth bones in central Brazil around the last glacial maximum. Proceedings of the Royal Society B: Biological Sciences, 290(2002). https://doi.org/10.1098/rspb.2023.0316
4. Llamas, B., Fehren-Schmitz, L., Valverde, G., Soubrier, J., Mallick, S., Rohland, N., Nordenfelt, S., Valdiosera, C., Richards, S. M., Rohrlach, A., Romero, M. I. B., Espinoza, I. F., Cagigao, E. T., Jiménez, L. W., Makowski, K., Reyna, I. S. L., Lory, J. M., Torrez, J. A. B., Rivera, M. A., & Burger, R. L. (2016). Ancient mitochondrial DNA provides high-resolution time scale of the peopling of the Americas. Science Advances, 2(4), e1501385. https://doi.org/10.1126/sciadv.1501385
5. Lesnek, A. J., Briner, J. P., Lindqvist, C., Baichtal, J. F., & Heaton, T. H. (2018). Deglaciation of the Pacific coastal corridor directly preceded the human colonization of the Americas. Science Advances, 4(5), eaar5040. https://doi.org/10.1126/sciadv.aar5040
6. Jonhson, J. (2016). Arlington man. National Parks Service. https://www.nps.gov/chis/learn/historyculture/arlington.htm
7. Grayson, D. K., & Meltzer, D. J. (2003). A requiem for North American overkill. Journal of Archaeological Science, 30(5), 585–593. https://doi.org/10.1016/s0305-4403(02)00205-4
8. Surovell, T. A., & Grund, B. S. (2012). The Associational Critique of Quaternary Overkill and why it is Largely Irrelevant to the Extinction Debate. American Antiquity, 77(4), 672–687. https://doi.org/10.7183/0002-7316.77.4.672
9. Fiedel, S., & Haynes, G. (2004). A premature burial: comments on Grayson and Meltzer’s “Requiem for overkill.” Journal of Archaeological Science, 31(1), 121–131. https://doi.org/10.1016/j.jas.2003.06.004
10. Surovell, T. A., & Waguespack, N. M. (2008). How many elephant kills are 14?: Clovis mammoth and mastodon kills in context. Quaternary International, 191(1), 82–97. https://doi.org/10.1016/j.quaint.2007.12.001
11. Grayson, D. K., & Meltzer, D. J. (2002). Clovis hunting and large mammal extinction: a critical review of the evidence. Journal of World Prehistory, 16, 313-359.
12. Waters, M. R., Stafford, T. W., Kooyman, B., & Hills, L. V. (2015). Late Pleistocene horse and camel hunting at the southern margin of the ice-free corridor: Reassessing the age of Wally’s Beach, Canada. Proceedings of the National Academy of Sciences, 112(14), 4263–4267. https://doi.org/10.1073/pnas.1420650112
13. Sanchez, G., Holliday, V. T., Gaines, E. P., Arroyo-Cabrales, J., Martinez-Taguena, N., Kowler, A., Lange, T., Hodgins, G. W. L., Mentzer, S. M., & Sanchez-Morales, I. (2014). Human (Clovis)-gomphothere (Cuvieronius sp.) association 13,390 calibrated yBP in Sonora, Mexico. Proceedings of the National Academy of Sciences, 111(30), 10972–10977. https://doi.org/10.1073/pnas.1404546111
14. María Elena Gutiérrez, Martínez, G. A., Luchsinger, H., Grill, S. C., Alejandro Fabián Zucol, Hassan, G. S., M. Teresa Barros, Kaufmann, C. A., & Calvo, I. (2011). Paleoenvironments in the Paso Otero locality during Late Pleistocene–Holocene (Pampean region, Argentina): An interdisciplinary approach. Quaternary International, 245(1), 37–47. https://doi.org/10.1016/j.quaint.2010.11.010
15. Politis, G. G., Messineo, P. G., Stafford, T. W., & Lindsey, E. L. (2019). Campo Laborde: A Late Pleistocene giant ground sloth kill and butchering site in the Pampas. Science Advances, 5(3), eaau4546. https://doi.org/10.1126/sciadv.aau4546
16. Moore, C., Kimball, L. R., Goodyear, A. C., Brooks, M. J., I. Randolph Daniel, West, A., Taylor, S., Weber, K., Fagan, J., & Walker, C. (2023). Paleoamerican exploitation of extinct megafauna revealed through immunological blood residue and microwear analysis, North and South Carolina, USA. Scientific Reports, 13(1). https://doi.org/10.1038/s41598-023-36617-z
17. Chatters, J. C., Potter, B. A., Fiedel, S. J., Morrow, J. E., Jass, C. N., & Wooller, M. J. (2024). Mammoth featured heavily in Western Clovis diet. Science Advances, 10(49). https://doi.org/10.1126/sciadv.adr3814
18. Ben-Dor, M., & Barkai, R. (2024). A matter of fat: Hunting preferences affected Pleistocene megafaunal extinctions and human evolution. Quaternary Science Reviews, 331, 108660. https://doi.org/10.1016/j.quascirev.2024.108660
19. Prates, L., & Perez, S. I. (2021). Late Pleistocene South American megafaunal extinctions associated with rise of Fishtail points and human population. Nature Communications, 12(1). https://doi.org/10.1038/s41467-021-22506-4
20. Martin, P. S., & Steadman, D. W. (1999). Prehistoric Extinctions on Islands and Continents. Springer EBooks, 17–55. https://doi.org/10.1007/978-1-4757-5202-1_2
21. Martin, P. S. (1973). The Discovery of America: The first Americans may have swept the Western Hemisphere and decimated its fauna within 1000 years. Science, 179(4077), 969–974. https://doi.org/10.1126/science.179.4077.969
22. Holdaway, R. N., Allentoft, M. E., Jacomb, C., Oskam, C. L., Beavan, N. R., & Bunce, M. (2014). An extremely low-density human population exterminated New Zealand moa. Nature Communications, 5(1). https://doi.org/10.1038/ncomms6436
23. Koch, P. L., & Barnosky, A. D. (2006). Late Quaternary Extinctions: State of the Debate. Annual Review of Ecology, Evolution, and Systematics, 37(1), 215–250. https://doi.org/10.1146/annurev.ecolsys.34.011802.132415
24. Waters, M. R., Keene, J. L., Forman, S. L., Prewitt, E. R., Carlson, D. L., & Wiederhold, J. E. (2018). Pre-Clovis projectile points at the Debra L. Friedkin site, Texas—Implications for the Late Pleistocene peopling of the Americas. Science Advances, 4(10), eaat4505. https://doi.org/10.1126/sciadv.aat4505
25. Hoffecker, J. F., Elias, S. A., Scott, G. R., O’Rourke, D. H., Hlusko, L. J., Potapova, O., Pitulko, V., Pavlova, E., Bourgeon, L., & Vachula, R. S. (2023). Beringia and the peopling of the Western Hemisphere. Proceedings of the Royal Society B: Biological Sciences, 290(1990). https://doi.org/10.1098/rspb.2022.2246
26. Estes, J. A., Burdin, A., & Doak, D. F. (2015). Sea otters, kelp forests, and the extinction of Steller’s sea cow. Proceedings of the National Academy of Sciences, 113(4), 880–885. https://doi.org/10.1073/pnas.1502552112
27. Haynes, G. (2013). Extinctions in North America’s Late Glacial landscapes. Quaternary International, 285, 89–98. https://doi.org/10.1016/j.quaint.2010.07.026
28. Metcalf, J. L., Turney, C., Barnett, R., Martin, F., Bray, S. C., Vilstrup, J. T., Orlando, L., Salas-Gismondi, R., Loponte, D., Medina, M., De Nigris, M., Civalero, T., Fernández, P. M., Gasco, A., Duran, V., Seymour, K. L., Otaola, C., Gil, A., Paunero, R., & Prevosti, F. J. (2016). Synergistic roles of climate warming and human occupation in Patagonian megafaunal extinctions during the Last Deglaciation. Science Advances, 2(6), e1501682. https://doi.org/10.1126/sciadv.1501682
29. Prates, L., & Perez, S. I. (2024). The Evidence for Human Agency in the Late Pleistocene Megafaunal Extinctions. Encyclopedia of the Anthropocene, 219–226. https://doi.org/10.1016/B978-0-443-14082-2.00035-1
30. Gómez Coutouly, Y. A., & Holmes, C. E. (2018). The microblade industry from Swan Point Cultural Zone 4b: Technological and cultural implications from the earliest human occupation in Alaska. American Antiquity, 83(4), 735–752. https://doi.org/10.1017/aaq.2018.38
31. Surovell, T. A., Pelton, S. R., Anderson-Sprecher, R., & Myers, A. D. (2015). Test of Martin’s overkill hypothesis using radiocarbon dates on extinct megafauna. Proceedings of the National Academy of Sciences, 113(4), 886–891. https://doi.org/10.1073/pnas.1504020112
32. Monteath, A. J., Gaglioti, B. V., Edwards, M. E., & Froese, D. (2021). Late Pleistocene shrub expansion preceded megafauna turnover and extinctions in eastern Beringia. Proceedings of the National Academy of Sciences, 118(52). https://doi.org/10.1073/pnas.2107977118
33. Prates, L., & Perez, S. I. (2021). Late Pleistocene South American megafaunal extinctions associated with rise of Fishtail points and human population. Nature Communications, 12(1). https://doi.org/10.1038/s41467-021-22506-4
34. R. Scott Byram, Lightfoot, K. G., & Jun Ueno Sunseri. (2024). Clovis points and foreshafts under braced weapon compression: Modeling Pleistocene megafauna encounters with a lithic pike. PLoS ONE, 19(8), e0307996–e0307996. https://doi.org/10.1371/journal.pone.0307996
35. Pires, M. M., Koch, P. L., Fariña, R. A., de Aguiar, M. A. M., dos Reis, S. F., & Guimarães, P. R. (2015). Pleistocene megafaunal interaction networks became more vulnerable after human arrival. Proceedings of the Royal Society B: Biological Sciences, 282(1814), 20151367. https://doi.org/10.1098/rspb.2015.1367
36. Owen-Smith, N. (1987). Pleistocene extinctions: the pivotal role of megaherbivores. Paleobiology, 13(3), 351–362. https://doi.org/10.1017/s0094837300008927
37. F. Robin O’Keefe, Dunn, R. E., Weitzel, E. M., Waters, M. R., Martinez, L., Binder, W. J., Southon, J., Cohen, J. E., Meachen, J., Larisa R.G. DeSantis, Kirby, M. E., Ghezzo, E., Brenner-Coltrain, J., Fuller, B. T., Farrell, A. B., Takeuchi, G. T., MacDonald, G. M., Davis, E. B., & Lindsey, E. (2023). Pre–Younger Dryas megafaunal extirpation at Rancho La Brea linked to fire-driven state shift. Science, 381(6659). https://doi.org/10.1126/science.abo3594
38. Gill, J. L., Williams, J. W., Jackson, S. T., Lininger, K. B., & Robinson, G. S. (2009). Pleistocene Megafaunal Collapse, Novel Plant Communities, and Enhanced Fire Regimes in North America. Science, 326(5956), 1100–1103. https://doi.org/10.1126/science.1179504
39. Raczka, M. F., Mosblech, N. A., Giosan, L., Valencia, B. G., Folcik, A. M., Kingston, M., Baskin, S., & Bush, M. B. (2019). A human role in Andean megafaunal extinction? Quaternary Science Reviews, 205, 154–165. https://doi.org/10.1016/j.quascirev.2018.12.005
40. G.M. Cárdenes-Sandí, C.R. Shadik, A. Correa-Metrio, Gosling, W. D., R. Cheddadi, & Bush, M. B. (2018). Central American climate and microrefugia: A view from the last interglacial. Quaternary Science Reviews, 205, 224–233. https://doi.org/10.1016/j.quascirev.2018.12.021
41. Schiferl, J., Kingston, M., Åkesson, C. M., Valencia, B. G., A. Rozas-Davila, McGee, D., Woods, A., Chen, C. Y., Hatfield, R. G., Rodbell, D. T., Abbott, M. B., & Bush, M. B. (2023). A neotropical perspective on the uniqueness of the Holocene among interglacials. Nature Communications, 14(1). https://doi.org/10.1038/s41467-023-43231-0
42. Schweger, C., Froese, D., White, J. M., & Westgate, J. A. (2011). Pre-glacial and interglacial pollen records over the last 3 Ma from northwest Canada: Why do Holocene forests differ from those of previous interglaciations? Quaternary Science Reviews, 30(17-18), 2124–2133. https://doi.org/10.1016/j.quascirev.2011.01.020
43. Miller, G. H. (2005). Ecosystem Collapse in Pleistocene Australia and a Human Role in Megafaunal Extinction. Science, 309(5732), 287–290. https://doi.org/10.1126/science.1111288
44. Mooney, S. D., Harrison, S. P., Bartlein, P. J., Daniau, A.-L. ., Stevenson, J., Brownlie, K. C., Buckman, S., Cupper, M., Luly, J., Black, M., Colhoun, E., D’Costa, D., Dodson, J., Haberle, S., Hope, G. S., Kershaw, P., Kenyon, C., McKenzie, M., & Williams, N. (2011). Late Quaternary fire regimes of Australasia. Quaternary Science Reviews, 30(1), 28–46. https://doi.org/10.1016/j.quascirev.2010.10.010
45. Johnson, C. N. (2016). Fire, people and ecosystem change in Pleistocene Australia. Australian Journal of Botany, 64(8), 643. https://doi.org/10.1071/bt16138
46. Barnosky, A. D., Lindsey, E. L., Villavicencio, N. A., Bostelmann, E., Hadly, E. A., Wanket, J., & Marshall, C. R. (2015). Variable impact of late-Quaternary megafaunal extinction in causing ecological state shifts in North and South America. Proceedings of the National Academy of Sciences, 113(4), 856–861. https://doi.org/10.1073/pnas.1505295112
47. Glover, K. C., George, J., Heusser, L., & MacDonald, G. M. (2021). West Coast vegetation shifts as a response to climate change over the past 130,000 years: geographic patterns and process from pollen data. Physical Geography, 1–19. https://doi.org/10.1080/02723646.2021.1990506
48 Pearce, E. A., Mazier, F., Normand, S., Fyfe, R., Andrieu, V., C.C. Bakels, Balwierz, Z., Krzysztof Bińka, Boreham, S., Borisova, O., Broström, A., Jacques‐Louis de Beaulieu, Gao, C., Penélope González-Sampériz, Wojciech Granoszewski, Hrynowiecka, A., Piotr Kołaczek, Petr Kuneš, Magri, D., & Malkiewicz, M. (2023). Substantial light woodland and open vegetation characterized the temperate forest biome before Homo sapiens. Science Advances, 9(45). https://doi.org/10.1126/sciadv.adi9135
49. Pearce, E. A., Mazier, F., Davison, C. W., Baines, O., Czyżewski, S., Fyfe, R., Bińka, K., Boreham, S., de Beaulieu, J.-L., Gao, C., Granoszewski, W., Hrynowiecka, A., Malkiewicz, M., Mighall, T., Noryśkiewicz, B., Pidek, I. A., Strahl, J., Winter, H., & Svenning, J.-C. (2025). Beyond the closed-forest paradigm: Cross-scale vegetation structure in temperate Europe before the late-Quaternary megafauna extinctions. Earth History and Biodiversity, 3, 100022. https://doi.org/10.1016/j.hisbio.2025.100022