On this blog, we’ve explored key aspects of the Pleistocene across multiple regions-South America, North America, Australia, and part of Asia. Today, we turn to Europe, a continent that has received perhaps the most attention in paleontological and Paleolithic research. The disproportionate wealth of information we have about Europe’s history is not only because of the richness of its fossil and archaeological record, but also because of its central role in the story of Western history and science.
Our focus will be on Europe during the Upper Paleolithic, a period which spanned roughly 50 thousand to 12 thousand years ago (the exact time designation depends on the authority and region). This era is particularly significant because it marks the first long-term establishment of our own species, Homo sapiens, on the continent. It was during this period that anatomically modern humans-known in Europe as Cro-Magnons or European Early Modern Humans-replaced their Neanderthal cousins. An explosion of art, such as cave paintings and figurines depicting living things, testifies to the creativity and vibrant social lives of these early representatives of our species while their survival in the face of harsh conditions speaks to their resilience and adaptability.
But this wasn’t just a human story, and we’ll begin by discussing the paleoclimatic, paleoenvironmental, and paleontological context of Europe before moving on to the lifestyles and origins of its prehistoric hunter-gatherers. Europe’s floral and faunal landscape were shaped by the profound glaciations of the last few million years. There was also a large and diverse community of animals, many of which no longer exist today. Understanding this ecological and environmental backdrop is essential to appreciating the lives of both humans and animals during this time.
And while the primary focus here will be the Upper Paleolithic in Europe, the information here will be placed within the broader context of the Quaternary period. In doing so, we’ll show how this pivotal chapter in Europe’s deep past connects to the broader story of changing environments and human history before and after the Upper Paleolithic.
This is a pretty long post and the amount of information might be overwhelming, so feel free to skip around to the parts that interest you the most. Having said that, let’s get right to today’s captivating topic.
Geographic and Ecological Background
Europe is usually considered a continent, although it’s often joined into the supercontinent “Eurasia” because it’s technically connected to Asia by land. Despite this connection, a number of geographic barriers such as the Urals, Caspian Sea, Black Sea, and Caucasus mountains separate Europe from Asia proper while the Mediterranean Sea separates Europe from both North Africa and Asia. Therefore, there are enough geographic barriers between Europe and the African and Asian continents to make Europe’s designation as its own continent seem reasonable, although it would probably make the most sense (in my opinion) to describe Europe as a subcontinent.
As expected for a landmass of its size, Europe has a range of different climates, but the climate of the majority of Europe can be described as mild. Southern Europe features Mediterranean climates with cool, rainy winters and hot, dry summers. A strong oceanic influence prevails over a wide swathe of western, central, and northern Europe with warm summers, cool winters, and precipitation that is fairly evenly spread out throughout the year. Winters are notably a lot warmer in Europe than at similar latitudes in Asia and North America due to the influence of the North Atlantic Current, a northern extension of the Gulf Stream. The climate becomes less oceanic the further east one travels with increasingly sharper contrasts between summer and winter temperatures. Still, the Black and Baltic seas help extend the maritime influence eastward.
Today, temperate broadleaf and mixed forests are the dominant biome in Europe, although temperate steppe and Mediterranean forest are also prominent. Taiga and tundra are restricted to the far north. When it comes to wildlife, red deer, roe deer, brown bear, wolves, wisent, ibex, and moose (called elk in Europe) comprise some of the last remaining large animals. More exotic animals such as Caucasian Turs and Persian leopards can be found in the Caucasus region at the very edge of Europe while saiga antelope can be found in the Eurasian steppe and reindeer in the far north. Many of these are critically endangered. The wild horse went extinct on the continent in the last few centuries, while the wisent had to be reintroduced into the wild recently after being eliminated in the wild.

However, during large parts of the Pleistocene, such as the Upper Paleolithic (UP), the climate and environment of Europe was alien compared to its present form. At various times, extensive ice sheets, polar deserts, tundra, and steppe stretched across what’s today lush forest while great herds of herbivores roamed the frozen wilderness as fierce predators pursued them. Let’s take a deeper look at the climatic, ecological, and anthropological context of this strange lost world.
Caught Between the Ice and Frozen Sea
As stated previously, Europe today has a relatively mild climate. However, this present state is not intrinsic: climates far different and less hospitable than those of today were the norm during the Pleistocene. Ice sheets repeatedly expanded and retreated over the past few million years, driven by Milankovitch cycles. During past glacial periods, Europe was dominated by two major ice sheets-the Alpine and the Fennoscandian. The Alpine glaciation covered the Alps while the Fennoscandian, at its largest extent, covered Scandinavia, much of the British Isles, the North European Plain, and parts of northwest Russia. The penultimate glaciation was even more extensive than the last one.
These ice sheets had complex and major impacts on local climate. They radically altered atmospheric circulation: the Fennoscandian ice sheet generated an anticyclone that drove winds from northeast to southwest, depositing vast amounts of loess (windblown dust) across the unglaciated parts of central Europe, while westerly storm tracks were displaced southward. However, a lot of the climatic peculiarities of ice age Europe were driven by the ocean, and specifically changes in oceanic circulation. To understand this, we’ll need to look into the system known as the Atlantic Meridional Overturning Circulation (AMOC) and what it does.
The AMOC is one of the most important features of Earth’s ocean dynamics. Normally, a current known as the Gulf Stream transports warm, salty tropical water northward along North America’s east coast and then toward western Europe, eventually releasing heat and moisture that moderate European winters (which would otherwise be a lot colder due to the latitude). This salty water becomes much colder and hence denser by the time it reaches the seas near Scandinavia, allowing it to sink to the bottom of the ocean. From there it flows back south, maintaining a “conveyor belt” circulation as more warm water flows northeastwards along the surface. The ocean west of Europe rarely freezes as warm water is continuously pumped along this circuit. Small correction: I originally described the AMOC bringing “relatively” fresh water northwards that then became saltier. In reality, the water is already quite salty as it leaves the subtropics.
However, this conveyor belt depends not only on a temperature gradient but also a salinity gradient. If the water is not salty enough by the time it reaches the northernmost seas, it will have a hard time sinking even if it’s become cold. As freshwater is less dense than saltwater, a large injection of the former can disrupt the AMOC process. And this occurred many times in the past: during glacial periods, massive freshwater inputs, such as from giant icebergs breaking off of ice sheets and melting in the ocean, periodically weakened or shut down the AMOC. Hence, warm tropical water did not flow effectively towards Europe, causing much cooler North Atlantic sea temperatures.
Sea temperatures were especially low in winter, causing extensive winter sea ice and rather cold temperatures over land during that season. Summer temperatures were more mildly impacted. Together with lower sea levels-which exposed areas like Doggerland and pushed the Atlantic westward-this reduced maritime influence and made much of Europe far more continental, resembling modern Siberia or interior Canada, with colder winters, drier conditions, and greater seasonal contrasts in temperature and precipitation. The combination of a colder ocean and ice sheets in northern Europe and the Alps helps explain how permafrost (frozen soil) expanded deep into central Europe during the coldest periods of glacials. See map below for reference:
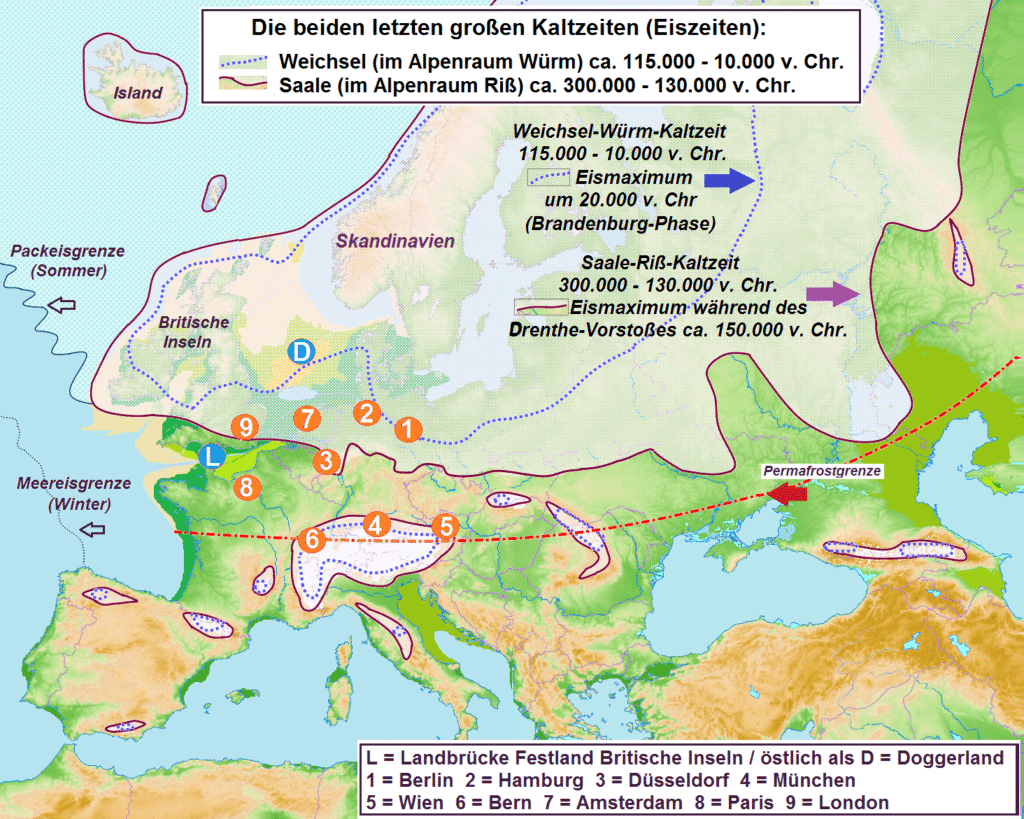
An important but oft-ignored factor was that CO2 levels were lower in glacial times, which further suppressed tree growth. These conditions could explain why Europe, which in recent times was dominated by forests, was instead primarily covered with steppe-tundra and forest-steppe vegetation. Nevertheless, the Pleistocene was not uniformly cold: there were of course stadials which represented the most quintessential “ice age” conditions but there were also interglacials like the Eemian which were as warm or warmer than today and intermediate states known as interstadials which were brief warm periods within glacials. Changes in oceanic circulation and shifting ice sheets also created an unstable climate that drove repeated range shifts among plants and animals as species expanded from and retreated into refugia.
Overall, Upper Paleolithic Europe was a harsh and volatile environment shaped by ice to the north and a turbulent ocean to the west. Still, this did not stop it from supporting a diverse and abundant assemblage of animals and vibrant human cultures.
Contrasts and Complexity In Paleofauna
I can recall taking a car from Charles De Gaulle airport to Paris in the summer of 2018. It was obvious to me from seeing the rich agricultural lands on the outskirts of the city that it’s an area with a pleasant climate. Major wine producing regions exist not far from the capital despite the location being at a similar latitude to cities in North America like Winnipeg or Quebec city. Therefore it feels jarring, almost inconceivable, when I hear that a tundra periodically covered France for much of prehistory.
Yet, this is reality. A vast cold tundra stretched across central Europe all the way to the Atlantic during ice ages. The technical name for this biome is steppe–tundra but it is also often called mammoth steppe. It had few trees. Low-growing plants from modern steppe and tundra biomes were instead dominant. It hosted famous bygone animals such as woolly mammoths, woolly rhinos, steppe bison, and cave lions who coexisted with extant creatures such as reindeer, saiga antelope, muskoxen, horses, and wolves.

Cave hyenas are either considered a subspecies of their African spotted hyena relatives or a closely related species. They were absent from the coldest parts of the mammoth steppe in northern Siberia but were indeed present in Europe where the climate was still somewhat milder. In addition to steppe-tundra, there were also forest-steppes which contained a larger proportion of trees and were home to many of the same animals, as well as the giant deer Megaloceros giganteus, also known as the Irish elk.
Lowered sea levels meant that rich grazing grounds on continental shelves were exposed, such as the Doggerland plain where the North Sea is currently located. Soils in large parts of Europe and northern Eurasia as a whole were enriched by the accumulation of glacially-deposited loess. Despite the short growing seasons, productive forage necessary to sustain large herds of animals was abundant. Data from the Eifel Maar in Germany indicates herbivores persisted through a variety of climatic changes over the past 60,000 years so long as vegetation remained open, with low concentrations associated with highly forested intervals.
It’s critical to note that not all animals associated with Pleistocene Europe were particularly adapted to cold and/or open habitats, however. There were also more temperate animals such as aurochs, red deer, wild boar, European straight-tusked elephants, hippos (first of the H. antiquus species, then the modern H. amphibius), roe deer, fallow deer, and narrow-nosed rhinos. The past presence of hippos in Europe is shocking in itself, but dholes, Barbary macaques, and a European relative of the water buffalo known as Bubalus murrensis lived there too.
These temperate creatures spread as far north as the British Isles during interglacials such as the Eemian but then retreated to various refugia in Europe during the deep cold of glacials, with the Mediterranean region being the most important refugium since it did not cool down as profoundly. These animals would have still been somewhat cold-tolerant because cool winter temperatures were unavoidable even in southern Europe during glacials, even if not to the same degree as in the steppe-tundra further north.
The Mediterranean is quite interesting because it actually hosted a number of species of dwarf proboscideans and hippopotami in its islands, descendants of mainland European relatives who became stranded there. Wisent, red deer, and wild boar are still present on the continent but the others have gone extinct locally (in Europe) or globally. The aurochs still technically has descendants in the form of the domestic cattle, however.

Two other extinct megafauna of Europe from this period included the famous cave bear and much less well-known Elasmotherium. The cave bear was a relative of the brown bear which was also present in Europe at the time and even seems to have interbred with it. It is considered to have been herbivorous but do not be mistaken in thinking it was a soft-target; it was known to injure and kill anything that tried to attack it from lions to humans. They were quite large, with males weighing up to 500 kg. It was generally associated with hilly areas rather than low lying mammoth steppe, possibly because these provide caves and probably had a lower density of predators.
Elasmotherium sibiricum was a rhinoceros that lived in the steppes of Eastern Europe and southern Siberia. It belonged to a lineage of rhinos, Elasmotheres, that diverged very early on from extant rhinos. Some have speculated that it had a very large horn (though this is disputed), giving it the name “Siberian unicorn”. Middle to Late Pleistocene E. sibiricum is believed to have been the last of the Elasmotheres.


Other quick mentions are three extant animals: wolves, moose, and leopards. Wolves in ice age Europe were part of a unique ecotype adapted to the mammoth steppe; they were replaced by modern wolves after the Last Glacial Maximum. Moose included the Early and Middle Pleistocene Cervalces latifrons (the massive broad-fronted moose), and its Late Pleistocene and Holocene descendant Alces alces , the modern moose. Although quite cold-hardy, moose avoided the mammoth steppe and preferred woodier areas. Leopards, which most people do not associate with Europe, actually once ranged as far west as Iberia. They are believed to have avoided the mammoth steppe and preferred more marginal habitat like hilly and/or wooded areas, likely to avoid larger and/or more dangerous predators. The site with the largest number of leopard remains (over 200) is Equi in the Italian Appenine mountains where remains of other predators are not common.

It is hard to overstate the extreme nature of faunal turnover in the European continent during the Quaternary. As stated earlier, animals like hippopotami and straight-tusked elephants extended into northwestern Europe during interglacials. Meanwhile, during glacials, reindeer, woolly mammoths, muskoxen, woolly rhinos, Arctic foxes, and wolverines could be found as far south as Spain and Italy (even southern Spain and Italy in some cases). It is especially remarkable because mountain ranges running east to west separate the Mediterranean peninsulas from central and northern Europe, complicating the dispersal ability of plants, animals, and even hominins in the face of warming or cooling.
This “changing of the guard” between glacial and temperate fauna happened multiple times in accordance with climate cycles. Given the especially rapid pace of climate change during glacial-interglacial transitions, it’s possible that in a given area, only a few thousand years may have separated the last occurrence of cold-adapted animals such as reindeer and mammoth on one hand and the first occurrence of relatively warm-adapted animals like hippopotami and straight-tusked elephants on the other.
Still, drawing a clean line between “glacial” and “temperate” assemblages, known as the Mammuthus-Coelodonta and Paleoloxodon faunal complexes respectively, is not necessarily easy. Some animals commonly associated with ice age Europe like Megaloceros also seem to have been widespread across the continent during interglacials. Cave hyenas had a broad range across Eurasia, including much of temperate Eurasia but also the southern edges of the mammoth steppe. There are also sites in Europe dating to the LGM where animals like woolly mammoths and woolly rhinos have been found alongside temperate animals like wild boar and red deer.


In fact, due to the presence of complex habitat mosaics-common then just as now-even animals with very different niches must have crossed paths. For example, woolly rhinos are known to have been cold-steppe grazers while moose are woodland browsers that also eat plenty of aquatic vegetation. Despite differences in habitat, it’s easy to imagine a woolly rhino entering a riverine wetland in search of water, where it encountered a moose browsing on shrubs. These kinds of intriguing but unexpected encounters between animals that preferred different habitats were likely more commonplace than thought. This is a fascinating thing to imagine.
Evidently, old Europe included a large number of creatures that are no longer with us, whether in the form of subspecies of existing animals or species in their own right. The causes of the extinctions are a matter of intense debate but I will not go into them in this particular article. It should be noted, however, that the extinctions did not take place all at once but rather over the course of tens of thousands of years from the start of the Upper Paleolithic to just the last few centuries.
Straight-tusked elephants, narrow-nosed rhinos, cave hyenas, Elasmotherium sibiricum, and hippopotami went extinct in Europe prior to the LGM, while cave bears went extinct around the LGM. Others such as cave lions, mammoths, and woolly rhinos disappeared from the continent between the LGM and the Holocene, while giant deer and European wild ass survived into the first half of the Holocene. Aurochs and tarpans (wild horses) survived into the late Holocene after a long period of decline. Future fossil discoveries may reveal that many of these animals survived longer than we thought. The staggered nature of extinctions in the European continent, and Eurasia as a whole, will be relevant in an upcoming article analyzing the causes of the Late Quaternary extinctions globally.
Regardless, the diverse and abundant array of animals served as the lifeline for the human hunter-gatherers of the Pleistocene, including Homo heidelbergensis, Neanderthals, and our own species. In large parts of Europe during the coldest phases, plant foods edible to humans were mostly absent, especially in winter. The histories of the animals and humans of this continent are deeply intertwined, and humans were just as profoundly affected by the environmental changes in Europe as the animals they relied on were. We will explore the intricacies of the lives of these humans, their interactions and relationships to each other, genomic histories, and more in the following sections.
The IUP: Neanderthals Out, Homo sapiens In
Perhaps the most famous inhabitants of Pleistocene Europe were the Neanderthals. They’re known from a number of sites across Europe (save for its northernmost parts which were covered by ice sheets or frozen tundra). The first Neanderthal specimen was Neander 1, named after the Neander valley in Rhineland, western Germany. Although human, they were part of what are known as archaic hominins and diverged from us-the anatomically modern humans-somewhere between 550 and 765 thousand years ago and are usually considered their own species: Homo neanderthalensis as opposed to Homo sapiens (us). Neanderthals descend from the earlier Homo heidelbergensis; both had a long history in the European continent spanning hundreds of thousands of years.
They were shorter and stockier with thicker bones and larger noses than anatomically modern humans (AMH); these physical traits were a necessary adaptation to help them deal with the cold. These hardy hominins were never very many in number and lived in small tight knit communities that seem to not have exchanged much with outsiders; the tools found in Neanderthal sites differed even from those in nearby locales which suggests that Neanderthal communities were independent and isolated to a large degree. In keeping with this trend, Neanderthal genomes also show a strong degree of inbreeding. Neanderthals, in many ways, may be likened to hermits.


The Middle Paleolithic Mousterian tool industry is associated with the Neanderthals. They are known to have hunted a variety of animals, including very large ones like elephants. However, it is likely that their dominant sustenance came from medium-sized animals like red deer. Many Neanderthals show signs of traumatic injuries resulting from intraspecies and inter-species conflict (mainly with predators such as bears and big cats); the rates of injuries resulting from violent encounters appears to be comparable to anatomically modern humans from the same time period.
While Neanderthals have been most closely associated with Europe in modern media and popular conception, they had a wide range stretching into southern Siberia, Central Asia, and West Asia (a region roughly corresponding to the Middle East). Indeed, leading Harvard geneticist David Reich argues that modern humans, including Europeans, do not possess DNA from European Neanderthals, but most likely their West Asian cousins. This speculation comes from the fact that all non-Africans seem to share similar amounts of Neanderthal ancestry that arrived in a single pulse around 50-55 kya (50,000-55,000 years ago), and somewhere in West Asia seems more likely than Europe because the former is directly in the path of a human population expanding out of Africa.
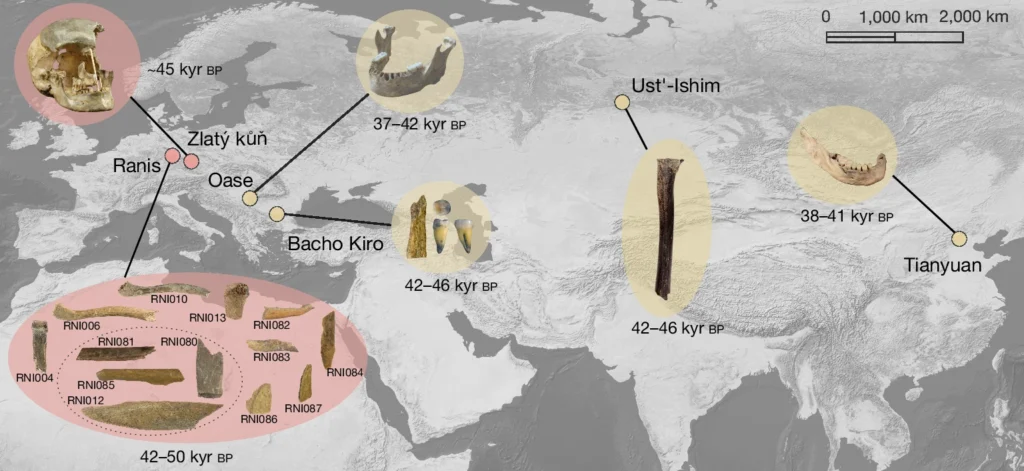
However, this does not mean that no humans interbred with Neanderthals in Europe. The pivotal period of human prehistory known as the Initial Upper Paleolithic (IUP) from 50 to 40 kya included the first confirmed expansion of Homo sapiens into Europe, where they encountered, admixed with, and seemingly replaced the Neanderthals. A few IUP specimens from Peștera cu Oase in Romania and Bacho Kiro in Bulgaria show evidence of recent interbreeding with Neanderthals in their lineage, with Neanderthal ancestry ranging from 3-6%, in excess of modern humans which are closer to 2%. Oase1 was dated to 42 to 37 kya (though this is probably an underestimate of its age) while the Bacho Kiro specimens lived between 46 and 42 kya.
Contemporaneous individuals from central Europe such as Zlatý kůň in the Czech Republic and Ranis in Germany appear to show no extra Neanderthal introgression compared to modern humans. It is possible that the ancestors of the Ranis and Zlatý kůň specimens encountered no or fewer Neanderthals compared to their Balkan counterparts. We may speculate on why this may have been the case-isotopic data from animal tooth enamel indicate both Ranis in Germany and Bacho Kiro in Bulgaria experienced very cold Subarctic climates during the initial period of human occupation. However, faunal remains from Bacho Kiro, unlike those from Ranis, contain some temperate elements, potentially suggesting climate heterogeneity in the former. It’s possible that more diverse conditions in the Balkans allowed local Neanderthals to hang on longer there than in east-central Europe.
No one really knows for sure how common modern human-Neanderthal interactions were, or what they were like. While it appears that Neanderthals were more intelligent and similar to humans than early anthropologists once believed, there do seem to be have been some noticeable cognitive differences between anatomically modern humans in Europe and the Neanderthals they encountered. AMH created art from UP Europe was on a level far beyond anything Neanderthals came up with. This is not to say that Neanderthals were less intelligent; it could just be that their cognitive strengths and weaknesses were distinct from our own. One possibility is that Neanderthals were innovative and intelligent but far too individualistic compared to humans, which may have given the more teamwork-oriented humans an advantage over them.
The fact that modern humans have Neanderthal ancestry, and that some extinct lineages had even more, would at first suggest that relations might have been amicable. Yet, it’s also possible that this interbreeding occurred under less pleasant, more violent circumstances as was unfortunately often the case throughout human history. We may never know the answer to that. We may also never know what factor, or combination of factors, was responsible for the ultimate demise of Neanderthals.
Being outcompeted by or even outright killed by AMH, deaths from AMH-introduced diseases, or being absorbed by the numerically superior AMH are all valid possibilities. Many have even argued climate change played a role, but the fact that Neanderthals disappeared from the entirety of their Eurasian range and not just Europe, as did at least two other archaic humans (Denisovans and Flores hobbits) following AMH expansion out of Africa could suggest the Neanderthal’s days were numbered regardless of what the climate did or did not do.
Regardless, the early European lineages of the IUP, including the ones that bred with European Neanderthals, were genetic dead ends (with the exception of Bacho Kiro that did give a trace of ancestry to later pops). They did not leave modern descendants and were themselves replaced by different, unrelated groups of anatomically modern humans. We will talk about those humans in the next section.
Cro Magnons: Artistic Cavemen
The term European Early Modern Human, or EEMH, is an umbrella term for humans living in Europe during the Upper Paleolithic, replacing the outdated (but cool sounding) term “Cro-Magnon”. These were not a single people; the IUPs as we just discussed went mostly extinct and had little to no connection with the groups that came later. The groups that followed were the Aurignacians, Gravettians, Solutreans, Magdalenians, and Epigravettians. The genetic relationships between the people of these cultures is a fascinating topic but it’s so complicated that I’ve decided to devote a separate, later section to it. However, it should quickly be noted that these peoples were genetically West Eurasian, which means they belonged to the same broad (key word here) clade of humanity as modern day Europeans and West Asians.
The Aurignacians were the earliest confirmed widespread EEMH culture in Europe (the validity of the allegedly earlier Chatelperronian is unconfirmed). Early people from the Goyet caves in Belgium are representatives of this culture, as well as possibly an individual from the Kostenki site in Russia. Aurignacians had diets that varied depending on the region, but in general they are strongly associated with reindeer which were possibly the most abundant medium-large animals in Europe at the time. In contrast to the flakes and thrusting weapons of the Neanderthal Mousterian industry, the Aurignacian featured blades and perhaps early projectiles (throwing weapons) for hunting.
The Aurignacian period is known for its extravagant artwork, such as the cave paintings at Chauvet in France, where a plethora of elegantly drawn animals are pictured next to each other. The exact purposes of cave paintings by UP humans are not known, but some have speculated they may have been the work of shamans drawing their visions. They also produced a number of exquisite sculptures of humans and animals. Among the most famous of these sculptures, made of mammoth ivory, was the “Lion-man” of Hohlestein-Stadel from Germany, which is exactly what it sounds like: it has the head of a cave lion and the body of a man.



The Gravettians followed the Aurignacians. They produced their own cave art and sculptures, just as the Aurignacians did. They are best known for their many figurines depicting plump women known as “Venuses”. Also prominent were engravings made on mammoth tusks. Gravettians also conducted elaborate burials, where the deceased were painted with red ochre and adorned with numerous grave goods such as beads, ivory, stone tools, and shells. The burials are known from central, eastern, and southern Europe but not western Europe, which could indicate that the Gravettians were not a culturally homogenous group.
Dolní Věstonice and Předmostí are the two most famous Gravettian sites, both located in Czechia in central Europe. Gravettians, especially those from central and eastern Europe, were strongly associated with mammoths. Countless mammoth bones have been found in and near Gravettian encampments in these regions, highlighting the importance of these creatures in the lives of the ancient people. Mammoths were frequently hunted for their meat, bones, and ivory. Structures were made from the bones. Gravettians of southern Europe probably took smaller game ranging from deer to hares as mammoths were fewer in number there.

Russia


A common theme so far, both among Neanderthals and the Cro-Magnons that followed, is the strong reliance on animal matter. This makes sense as plant foods edible to hominins were in short supply in this cold climate, particularly in winter. But relying heavily on animals was not without challenges. Small animals could only provide so much sustenance because they contain little fat, and excessive protein intake can be dangerous. Therefore, humans were highly dependent on large herbivores-which contained a lot more fat-in order to meet their nutritional needs, and declines in the latter’s populations could have proven fatal.
Furthermore, humans would have needed a way to obtain sufficient amounts of Vitamin C despite the lack of plants in order to avoid scurvy. For coastal populations, this may not have been as much of an issue because marine animals and seaweed contain plenty of Vitamin C, but inland populations would have a harder time. It is possible that it was obtained from the organs of animals but even then, the supply of Vitamin C could have been erratic and been a limiting factor for human groups.
Moving on, deteriorating conditions heading into the Last Glacial Maximum (LGM) around 23 kya caused humans to abandon central Europe and the reign of the Gravettians to end as ice sheets and polar deserts reached their maximum extents. Central Europe was undoubtedly in the grip of severe cold during the LGM, but paleoclimate simulations do not indicate that it was colder than places in the Arctic or Subarctic in recent times, where groups such as Yakuts and Inuits thrived long before modern housing and heating were invented. In fact, humans were in the Arctic circle in northeastern Siberia (Yana) 32,000 years ago where winter temperatures should have been even colder than in LGM Europe, which made me personally wonder if factors beyond pure cold were responsible for humans being unable to survive in the latter.
One study may provide an answer. Wren and Burke (2019) speculate that climate variability was critical in determining the distribution of humans in Europe. The authors of the study distinguish climate variability, which refers to annual and seasonal variation, from climate change which works on longer timescales; a high degree of variability means that the abundance and distribution of resources (animals, water, etc.) will fluctuate unpredictably which is bad for hominins who seek to minimize risk. Large areas of unglaciated Europe apparently had high climate variability and therefore lacked suitable habitat.
Regardless, not all of Europe was inhospitable for humans even during the LGM. The Franco-Cantabrian region of southern France and northern Spain was the most important refuge for humans during the Upper Paleolithic, including during the harsh LGM conditions. Today, this incredibly scenic part of the continent is located in Basque country, and it features mountain ranges and highlands such as the Cantabrians, the Pyrenees, and the Massif Central. Located next to the Bay of Biscay, the climate is strongly oceanic with plentiful rainfall which is spread out evenly by season in contrast to the Mediterranean climates further south which feature summer droughts. The Cantabrian area of northern Spain, also called “Green Spain”, is known for its lush forests.

The pleasant conditions extended back into the Pleistocene. Adequate rainfall, mild temperatures (though cooler than at present), access to the sea, and varied topography made the area resource-rich and hence appealing to hunter gatherers. It is not a coincidence that the area is dense in archeological sites, including ones that gave their names to the prehistoric cultures such as Aurignac, Gravette, Solutre, and La Madeleine, as well as caves like Lascaux, Chauvet, El Castillo, and Altamira displaying magnificent art. In fact, even the name Cro-Magnon is derived from a rock shelter in southwestern France.
Solutreans emerged in the Franco-Cantabrian refuge from a western population of Gravettians who were fleeing the deteriorating conditions further north. They exploited a variety of foods in this resource-rich region with red deer and ibex being amongst the most important game. To the east of the Solutreans were the Epigravettians of the Italian peninsula, Balkans, and the Pontic-Caspian steppe but despite the name, Epigravettians do not descend from Gravettians. The Solutreans and Epigravettians were separated from each other during the LGM by the glaciation of the Alps.
As the LGM ended, the Solutrean gave way to the Magdalenian around 17,000 years ago, or possibly a bit earlier. It is believed that the paintings at Lascaux were the work of the Magdalenians. These individuals were horse and reindeer hunters who appear to have made use of atlatls (spear-throwers). They expanded from the Franco-Cantabrian refuge into northern France, Germany, and Poland as the climate warmed. Meanwhile, the Epigravettians burst out of their own refuges in the Italian and Balkan peninsulas and expanded deep into the rest of Europe.
The period following the Paleolithic, the Mesolithic, was characterized by the emergence of a number of hunter-gatherer cultures in different parts of Europe such as the Azilian, Iron Gates, Komsa, Neman, and many others. The period as a whole encompassed the late glacial period to the middle of the Holocene. However, the Mesolithic lasted longer in northern Europe than in southern Europe as agriculture-made possible by the stable and warm conditions of the Holocene-first spread to the Mediterranean from the Near East.
Megafauna were not as plentiful during the Mesolithic as they were during the Paleolithic, as many species had disappeared but there were still animals like red deer, wild boar, aurochs, moose, and wisent which were enough to sustain the foragers. However, the dense forests that had overtaken large parts of Europe starting from the Bølling–Allerød warming onwards probably meant that hunting strategies were different from earlier, more open periods. Additionally, the climatic amelioration would have allowed wild plant foods to flourish; indeed, various types of nuts (especially hazelnuts) and berries were clearly important in the diets of these peoples.
We must remember that while the lives of these prehistoric peoples were fascinating and rich, they were not exactly peaceful. They would have sustained injuries or died in the process of hunting, and violent conflicts between individuals and groups appear to have been common just as they were in all human societies. There is evidence of cannibalism being practiced among both Neanderthals and later Cro-Magnons (especially Magdalenians). But having briefly acknowledged this sobering reality, we can move on to exploring the intriguing relationships between these groups.
Diving Deeper Into Hunter-Gatherer Genetics
We have already spoken earlier about the genetics of Neanderthals, so we’ll start off with their immediate successors, the IUPs. IUPs were a diverse group; Zlatý kůň and Ranis from central Europe appear to have belonged to lineages that split from Eurasians prior to the split between East and West Eurasians. Bacho Kiro and Oase1 from the Balkans had elevated Neanderthal ancestry and show more affinity to modern East Asians than to Europeans, and the same is true of Ust’-Ishim man from Siberia. Aside from Bacho Kiro individuals who appear to have contributed to the later Aurignacians, these lineages went extinct and are highly distinct from later Cro-Magnons as well as modern Europeans.
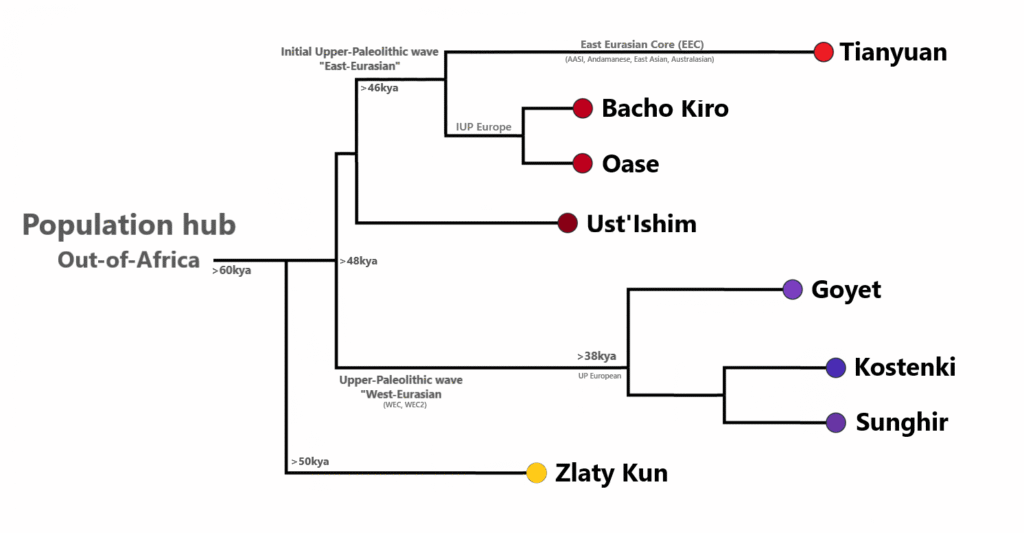
The Aurignacians that came later are best represented by early individuals from the Goyet cave. An individual “Q116-1” from Goyet appears to have minor Bacho Kiro IUP-like ancestry, but they and other Aurignacian individuals still appear to be primarily West Eurasian. The cultural affiliation of a 37 kya individual from the Kostenki site of Russia, Kostenki-14, is not clear; he may be part of an eastern branch of Aurignacians or perhaps something else altogether. He is most closely related to later Gravettian individuals from the same site as well as to Sunghir, also in Russia.
Western and southwestern Gravettians belonging to the Fournol cluster-named after the Fournol site in southern France-descend from Aurignacian individuals from Goyet in Belgium, with some geographically intermediate individuals falling in between Vestonice and Fournol genetically. Gravettians from eastern and southern Europe belong to what is known as the Vestonice cluster, named after the Dolní Věstonice type-site, and appear to have affinities with the pre-30 kya Kostenki and Sunghir individuals from Russia. A preprint by Gelabert et al. (2025) suggests that Sunghir-type ancestry also made it into some southwestern Gravettians via analysis of the genome of a 30 kya old female from Isturitz in southwestern France, showing that population exchanges between east and west were complex.
The Solutreans were founded by Gravettians belonging to the Fournol cluster, which again were descended from Aurignacians from Goyet such as Q116-1. The Epigravettians, represented by the Villabruna cluster, do not seem to be closely related to western or eastern Gravettians but instead had affinities to modern and ancient Near Eastern populations. After the LGM, Epigravettians spread over a large swathe of Europe. The Magdalenians also spread out across much of central Europe but were absorbed by Epigravettians. Actually, the Magdalenians themselves already had Villabruna-type ancestry in addition to their Solutrean/Fournol roots-the mixture represented by the “Goyet Q2 cluster”. The Oberkassel cluster was created by the full absorption of the Magdalenians by the Epigravettians.
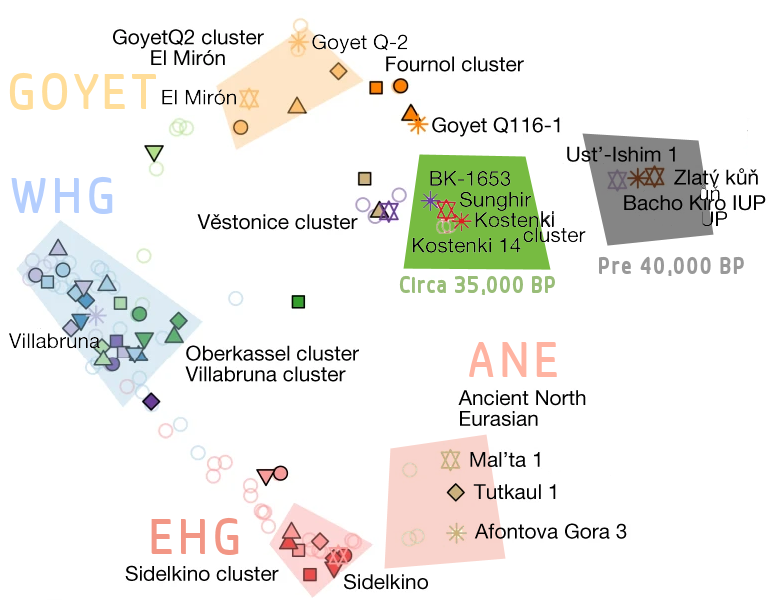
The Oberkassel cluster is synonymous with the Western European Hunter Gatherers (WHG) who dominated a broad swathe of Europe through the Mesolithic and were represented by cultures such as the Azilian in western Europe. The Eastern European Hunter Gatherers (EHG) to the east of the WHG were mostly descended from a distinct population from further east in Siberia known as Ancient North Eurasians. These, along with Scandinavian hunter gatherers, were the dominant genetic groupings of Mesolithic Europe, and mixed EHG-WHG populations could be found in places like the Balkans, Ukraine, and the Baltics.
The appearance of WHG has been a subject of controversy but it is believed through genotyping that they were blue-eyed with dark skin relative to modern Europeans, as seen in the reconstruction below. Physical reconstructions of Aurignacians and Gravettians also usually depict tan skin tones. Cheddar Man from the UK and Oberkassel from Germany are notable representatives of this group. EHGs, meanwhile, seem to have been pale with dark eyes. Sidelkino and Samara hunter gatherers are notable EHGs from Russia.


A consistent theme here is population replacement, and no strong genetic continuity. The Neanderthals disappear, as do the IUPs. Central and eastern Gravettians disappear during the LGM and leave little trace. The Magdalenians overwhelmingly get absorbed by Epigravettian types. And interestingly, a less extreme version of this trend continued well into the Holocene. Mesolithic WHG, themselves mainly derivative of Epigravettians who arrived at the end of the LGM, only contribute a modest amount to the modern European genome, with much larger proportions devoted by Neolithic Anatolian Farmers and Bronze Age Steppe-Herders (mixes between Eastern European and Caucasus Hunter Gatherers) who absorbed them. Given all these genetic transformations, we can safely say that pre-LGM Europeans like Aurignacians and Gravettians contributed only traces of ancestry to modern Europeans.
However, Holocene European migrations involved resident groups being assimilated by people with vastly different lifestyles, hunter gatherers were absorbed by farmers, who were then absorbed by steppe herders. Paleolithic demographic changes exclusively involved hunter-gatherer groups and sometimes included overwhelming replacement or even extinction of ancestries. This may indicate that the causes of population shifts were different between the Late Pleistocene and Holocene, with climate change possibly being more important in the former and cultural changes more in the latter.
To those not already familiar with European paleogenetics, this information may seem ridiculously confusing (because it is). Yet, this is the nature of demographic change over time, especially but not exclusively in Europe. While it is undeniably true that the modern peoples of Europe descend from people who settled the continent many thousands of years ago, aDNA has shown us that the picture becomes more complex beyond this. Modern Europeans descend from Holocene pulses of admixture between disparate groups, whereas many cultures from UP Europe left little or no modern ancestry. It was once thought that today’s people descend directly from their ancestors who first settled their respective regions tens of thousand of years in the past-Europe is a great place to show how human history usually defies this neat and simplistic notion.
Final Thoughts
All in all, the Upper Paleolithic in Europe stands out because it could not have been any more different from the Europe of today. The climate and environment were essentially alien to the modern day, and there was whole host of animals many of which are no longer present in Europe or even the rest of the world. It was this unrecognizable setting the first members of our species encountered when they settled the continent.
Both the first Homo sapiens and the Neanderthals that preceded them had to cope with the typical challenges of of hunter-gatherer life but in addition to problems more specific to those living in particular climate zones such as intense cold and a scarcity of plant foods. Not only was the climate far harsher than today, it was also subject to powerful changes which strongly affected the humans of the time just as it had affected their animal counterparts throughout the Pleistocene. Climate change seems to have played a major role in migrations and population replacement among the prehistoric humans, which explains the genomic evidence indicating relatively low continuity between earlier Upper Paleolithic groups and those present by the Mesolithic.
Europe has been studied more than anywhere else, and we owe our relatively deep understanding of the environment and human cultures of its prehistory to this fact. But ongoing research elsewhere is starting to close the gap, and it will be fascinating to see how complex the story of paleoecology and archeology in other parts of the world ends up being as the picture becomes clearer.
References
Álvarez-Lao, D. J., & García, N. (2011). Geographical distribution of Pleistocene cold-adapted large mammal faunas in the Iberian Peninsula. Quaternary International, 233(2), 159–170. https://doi.org/10.1016/j.quaint.2010.04.017
Barlow, A., Cahill, J. A., Hartmann, S., Theunert, C., Xenikoudakis, G., Fortes, G. G., Paijmans, J. L. A., Rabeder, G., Frischauf, C., Grandal-d’Anglade, A., García-Vázquez, A., Murtskhvaladze, M., Saarma, U., Anijalg, P., Skrbinšek, T., Bertorelle, G., Gasparian, B., Bar-Oz, G., Pinhasi, R., & Slatkin, M. (2018). Partial genomic survival of cave bears in living brown bears. Nature Ecology & Evolution, 2(10), 1563–1570. https://doi.org/10.1038/s41559-018-0654-8
Beier, J., Anthes, N., Wahl, J., & Harvati, K. (2018). Similar cranial trauma prevalence among Neanderthals and Upper Palaeolithic modern humans. Nature, 563(7733), 686–690. https://doi.org/10.1038/s41586-018-0696-8
Bosq, M., Kreutzer, S., Bertran, P., Lanos, P., Dufresne, P., & Schmidt, C. (2023). Last Glacial loess in Europe: luminescence database and chronology of deposition. Earth System Science Data, 15(10), 4689–4711. https://doi.org/10.5194/essd-15-4689-2023
Dale Guthrie, R. (2001). Origin and causes of the mammoth steppe: a story of cloud cover, woolly mammal tooth pits, buckles, and inside-out Beringia. Quaternary Science Reviews, 20(1-3), 549–574. https://doi.org/10.1016/s0277-3791(00)00099-8
Gaudzinski-Windheuser, S., Kindler, L., & Wil Roebroeks. (2023). Widespread evidence for elephant exploitation by Last Interglacial Neanderthals on the North European plain. Proceedings of the National Academy of Sciences of the United States of America, 120(50). https://doi.org/10.1073/pnas.2309427120
Gelabert, P., Sawyer, S., Cheronet, O., Villalba-Mouco, V., Oberreiter, V., Gonzalez-Morales, M. R., Straus, L. G., Igor Gutierrez-Zugasti, Cuenca-Solana, D., Garate, D., Marin-Arroyo, A. B., Lumley, H. de, Tejero, J.-M., Normand, C., Darricau, J., Lucci, M., Nava, A., Genchi, F., Coppola, D., & Marca, C. L. (2025). Genetic admixture between East and West European Gravettian-associated populations in Western Europe before the Last Glacial Maximum. BioRxiv (Cold Spring Harbor Laboratory). https://doi.org/10.1101/2025.09.26.678744
Ghezzo, E., & Rook, L. (2015). The remarkable Panthera pardus (Felidae, Mammalia) record from Equi (Massa, Italy): taphonomy, morphology, and paleoecology. Quaternary Science Reviews, 110, 131–151. https://doi.org/10.1016/j.quascirev.2014.12.020
Günther, T., Malmström, H., Svensson, E. M., Omrak, A., Sánchez-Quinto, F., Kılınç, G. M., Krzewińska, M., Eriksson, G., Fraser, M., Edlund, H., Munters, A. R., Coutinho, A., Simões, L. G., Vicente, M., Sjölander, A., Jansen Sellevold, B., Jørgensen, R., Claes, P., Shriver, M. D., & Valdiosera, C. (2018). Population genomics of Mesolithic Scandinavia: Investigating early postglacial migration routes and high-latitude adaptation. PLOS Biology, 16(1), e2003703. https://doi.org/10.1371/journal.pbio.2003703
Hajdinjak, M., Mafessoni, F., Skov, L., Vernot, B., Hübner, A., Fu, Q., Essel, E., Nagel, S., Nickel, B., Richter, J., Moldovan, O. T., Constantin, S., Endarova, E., Zahariev, N., Spasov, R., Welker, F., Smith, G. M., Sinet-Mathiot, V., Paskulin, L., & Fewlass, H. (2021). Initial Upper Palaeolithic humans in Europe had recent Neanderthal ancestry. Nature, 592(7853), 253–257. https://doi.org/10.1038/s41586-021-03335-3
Helmens, K. F. (2014). The Last Interglacial–Glacial cycle (MIS 5–2) re-examined based on long proxy records from central and northern Europe. Quaternary Science Reviews, 86, 115–143. https://doi.org/10.1016/j.quascirev.2013.12.012
José Luis Guil‐Guerrero. (2023). The Role of Large Mammals as Vitamin C Sources for MIS 3 Hominins. Quaternary, 6(1), 20–20. https://doi.org/10.3390/quat6010020
Kosintsev, P., Mitchell, K. J., Devièse, T., van der Plicht, J., Kuitems, M., Petrova, E., Tikhonov, A., Higham, T., Comeskey, D., Turney, C., Cooper, A., van Kolfschoten, T., Stuart, A. J., & Lister, A. M. (2018). Evolution and extinction of the giant rhinoceros Elasmotherium sibiricum sheds light on late Quaternary megafaunal extinctions. Nature Ecology & Evolution, 3(1), 31–38. https://doi.org/10.1038/s41559-018-0722-0
Lister, A. M., & Stuart, A. J. (2019). The extinction of the giant deer Megaloceros giganteus (Blumenbach): New radiocarbon evidence. Quaternary International, 500, 185–203. https://doi.org/10.1016/j.quaint.2019.03.025
Marsh, W. A., & Bello, S. (2023). Cannibalism and burial in the late Upper Palaeolithic: Combining archaeological and genetic evidence. Quaternary Science Reviews, 319, 108309. https://doi.org/10.1016/j.quascirev.2023.108309
Mol, D., & Bakker, R. (2022). Quaternary terrestrial megafaunal remains and their localities in the southern bight of the North Sea between the British Isles and The Netherlands: An overview. Staringia, 17(1), 88–127. https://natuurtijdschriften.nl/pub/1024370
Pacher, M., & Stuart, A. J. (2009). Extinction chronology and palaeobiology of the cave bear (Ursus spelaeus). Boreas, 38(2), 189–206. https://doi.org/10.1111/j.1502-3885.2008.00071.x
Pederzani, S., Britton, K., Aldeias, V., Bourgon, N., Fewlass, H., Lauer, T., McPherron, S. P., Rezek, Z., Sirakov, N., Smith, G. M., Spasov, R., Tran, N.-H., Tsanova, T., & Hublin, J.-J. (2021). Subarctic climate for the earliest Homo sapiens in Europe. Science Advances, 7(39). https://doi.org/10.1126/sciadv.abi4642
Pederzani, S., Britton, K., Trost, M., Fewlass, H., Bourgon, N., McCormack, J., Klervia Jaouen, Dietl, H., Hans-Jürgen Döhle, Kirchner, A., Lauer, T., Mael Le Corre, McPherron, S. P., Meller, H., Mylopotamitaki, D., Jörg Orschiedt, Rougier, H., Ruebens, K., Schüler, T., & Virginie Sinet-Mathiot. (2024). Stable isotopes show Homo sapiens dispersed into cold steppes ~45,000 years ago at Ilsenhöhle in Ranis, Germany. Nature Ecology and Evolution. https://doi.org/10.1038/s41559-023-02318-z
Posth, C., Yu, H., Ghalichi, A., Rougier, H., Crevecoeur, I., Huang, Y., Ringbauer, H., Rohrlach, A. B., Nägele, K., Villalba-Mouco, V., Radzeviciute, R., Ferraz, T., Stoessel, A., Tukhbatova, R., Drucker, D. G., Lari, M., Modi, A., Vai, S., Saupe, T., & Scheib, C. L. (2023). Palaeogenomics of Upper Palaeolithic to Neolithic European hunter-gatherers. Nature, 615(7950), 117–126. https://doi.org/10.1038/s41586-023-05726-0
Prüfer, K., Racimo, F., Patterson, N., Jay, F., Sankararaman, S., Sawyer, S., Heinze, A., Renaud, G., Sudmant, P. H., de Filippo, C., Li, H., Mallick, S., Dannemann, M., Fu, Q., Kircher, M., Kuhlwilm, M., Lachmann, M., Meyer, M., Ongyerth, M., & Siebauer, M. (2013). The complete genome sequence of a Neanderthal from the Altai Mountains. Nature, 505(7481), 43–49. https://doi.org/10.1038/nature12886
Reich, D. (2019). Who we are and how we got here : ancient DNA and the new science of the human past. Oxford University Press.
Rustioni, M., Ferretti, M. P., Mazza, P., Pavia, M., & Varola, A. (2003). The vertebrate fauna from Cardamone (Apulia, southern Italy): an example of Mediterranean mammoth fauna. Deinsea, 9(1), 395–404. https://natuurtijdschriften.nl/pub/538693
Sarunas Milisauskas. (2012). European Prehistory. Springer Science & Business Media.
Sirocko, F., Albert, J., Britzius, S., Dreher, F., Martínez-García, A., Dosseto, A., Burger, J., Terberger, T., & Haug, G. (2022). Thresholds for the presence of glacial megafauna in central Europe during the last 60,000 years. Scientific Reports, 12(1), 20055. https://doi.org/10.1038/s41598-022-22464-x
Slimak, L., Tharsika Vimala, Andaine Seguin-Orlando, Metz, L., Zanolli, C., Renaud Joannes-Boyau, Frouin, M., Arnold, L. J., Demuro, M., Thibaut Devièse, Comeskey, D., Buckley, M., Camus, H., Muth, X., Lewis, J. E., Hervé Bocherens, Pascale Yvorra, Christophe Tenailleau, Duployer, B., & Hélène Coqueugniot. (2024). Long genetic and social isolation in Neanderthals before their extinction. Cell Genomics, 4(9), 100593–100593. https://doi.org/10.1016/j.xgen.2024.100593
Sommer, R. S., & Nadachowski, A. (2006). Glacial refugia of mammals in Europe: evidence from fossil records. Mammal Review, 36(4), 251–265. https://doi.org/10.1111/j.1365-2907.2006.00093.x
Strandberg, G., Brandefelt, J., Kjellstro¨M, E., & Smith, B. (2011). High-resolution regional simulation of last glacial maximum climate in Europe. Tellus A: Dynamic Meteorology and Oceanography, 63(1), 107–125. https://doi.org/10.1111/j.1600-0870.2010.00485.x
Stuart, A. J., & Lister, A. M. (2007). Patterns of Late Quaternary megafaunal extinctions in Europe and northern Asia.
Sümer, A. P., Rougier, H., Villalba-Mouco, V., Huang, Y., Leonardo, Essel, E., Mesa, A. B., Furtwaengler, A., Stéphane Peyrégne, Filippo, C. de, Rohrlach, A. B., Pierini, F., Fabrizio Mafessoni, Fewlass, H., Zavala, E. I., Mylopotamitaki, D., Bianco, R. A., Schmidt, A., Zorn, J., & Nickel, B. (2024). Earliest modern human genomes constrain timing of Neanderthal admixture. Nature. https://doi.org/10.1038/s41586-024-08420-x
Vallini, L., Marciani, G., Aneli, S., Bortolini, E., Benazzi, S., Pievani, T., & Pagani, L. (2022). Genetics and Material Culture Support Repeated Expansions into Paleolithic Eurasia from a Population Hub Out of Africa. Genome Biology and Evolution, 14(4). https://doi.org/10.1093/gbe/evac045
Wren, C. D., & Burke, A. (2019). Habitat suitability and the genetic structure of human populations during the Last Glacial Maximum (LGM) in Western Europe. PLOS ONE, 14(6), e0217996. https://doi.org/10.1371/journal.pone.0217996
Wynn, T., Overmann, K., & Coolidge, F. (2016). The false dichotomy: a refutation of the Neandertal indistinguishability claim. JASs Invited Reviews Journal of Anthropological Sciences the JASs Is Published by the Istituto Italiano Di Antropologia Www.isita-Org.com, 94, 201–221. https://doi.org/10.4436/jass.94022
Zander, P. D., Böhl, D., Sirocko, F., Auderset, A., Haug, G. H., & Martínez-García, A. (2024). Reconstruction of warm-season temperatures in central Europe during the past 60 000 years from lacustrine branched glycerol dialkyl glycerol tetraethers (brGDGTs). Climate of the Past, 20(4), 841–864. https://doi.org/10.5194/cp-20-841-2024