Pleistocene Ice Ages conjure up notions of intense cold, massive glaciers, and an endless wilderness hosting a large and diverse community of animals. Most people understand that it was a very different world. Yet, if a layman were to go tens of thousands of years back in time, the landscapes they would encounter would not appear noticeably different to those that can be found in our planet today, save for a much larger biomass and diversity of large animals. One could easily make out familiar ecosystems such as forests, steppes, tundras, deserts, and so on, albeit shifted geographically due to differences in climate.
However, to a person well versed in ecology, glacial plant and animal communities would appear rather peculiar as they were often distinct in terms of their species composition when compared to any modern ecosystems. These are what are known as no-analog communities. To be clear, this does not refer to the past flora and fauna of particular regions differing from their respective present-day flora and fauna (which they also did), but rather their difference from the particular mix of flora and fauna found anywhere today. Proportions matter too; for example, even if the same plants occurred in a glacial ecosystem as in a certain modern ecosystem, the relative abundance of the taxa may diverge significantly.
I would like to provide a short disclaimer before going further: I am far from an expert in botany and have quite a lot to learn when it comes to plants, but I nonetheless can understand and appreciate the uniqueness of glacial plant communities from what I’ve read. With that said, we can move on to exploring these fascinating communities without modern matches.
No-analog Plant Communities
A massive ice sheet known as the Laurentide Ice Sheet covered the northern half of the eastern United States during the Wisconsin glaciation. A narrow band of tundra was situated directly south of the ice sheet, which then gave way to forests that vaguely resembled today’s boreal forests situated much further north. At the edge of this tundra-forest boundary in the Lake Erie region, high amounts of pine pollen has been found alongside pollen of tundra plants from 55 to 50 thousand radiocarbon years before present (somewhat earlier in calendar years)1. This is surprising and indicates some kind of forest-tundra ecosystem where the trees were heavily pine. Such an ecosystem no longer exists because the conifers at the southern boundary of modern tundras tend to be spruces.
The forests south of the ice sheet were not quite same as the modern boreal forests of Alaska or Canada; they tended to be more open with an abundance of herbaceous vegetation such as grasses, sedges, and forbs, and might be better described as woodlands or parklands. In the Ozarks of western Missouri, a parkland consisting of pine and oaks existed between 40 and 25 thousand radiocarbon years BP1. This has been compared to ecotones (transitional ecosystems) such as the mixed pine-hardwood forest of northwestern Minnesota, but the pine in the Ozarks was jack pine rather than white pine. It has also been compared to the aspen parkland in Manitoba but was different in having a lower proportion of spruce amongst the trees and high representation of sedges among the herbs.

Shifting now from forests to open landscapes, a vast biome known as the mammoth steppe, or steppe-tundra, prevailed across large swathes of the Holarctic2. It is perhaps the best known of glacial ecosystems with few analogs, containing a mixture of plant and animal species that are rarely found together today. During glacial periods, higher than present aridity and continentality caused the forest belt of northern Eurasia to disappear3, possibly in conjunction with low CO2 which suppressed woody vegetation. The Arctic tundra belt shifted south and the Eurasian steppe zone shifted north and both met in the middle, causing their respective plant and animal taxa to mix and mingle along a very broad front from western Europe to the Yukon. Hence, “steppe-tundra”.
This biome was not exactly homogenous and probably included a mosaic of habitats, some dominated by dry steppe and others by moist-tundra plants4. So while it contained a mixture of plant types from steppe and tundra, it was not necessarily a thorough blend. There actually is something of an analog to this in the southern Siberian Altai (which I have written about previously), where the varied topography and intersection of different vegetation zones creates a modern relic of this lost biome5.

As with plant communities, animal communities differed in their compositions as well. Today, saiga antelope and reindeer are not usually found together, with the former occupying temperate steppes and the latter being present in tundra and taiga, but the two shared a wide swathe of territory during the Pleistocene. This is because the no-analog mammoth steppe met both of their requirements, with plentiful grasses and low snow cover supporting the saiga and the low temperature and presence of mosses and lichen supporting reindeer. Just like plants, animals have their own unique requirements for survival and conditions could have been sufficiently distinct in the past to accommodate different mixtures of animal.
Causes of Unusual Associations
The causes for the dissimilarity between glacial and modern pollen assemblages are debated. The steppe-tundra may be explained by the fact that the decimation of the north Eurasian forest belt allowed for a diffusion and mingling of steppe and tundra plants, but the no-analog forests of North America are harder to explain. Lower CO2 and the presence of megafauna could have contributed to the relative openness of these forests (which again might be better described as parkland or woodland) but the unusual proportions of tree types relative to each other probably had different primary causes6.
In eastern North America, much research on the topic has centered on vegetation dynamics during the last deglaciation period where extensive no-analog forests existed during a time of rapid climate change and high continentality. No-analog vegetation was more widespread during this period than during the Last Glacial Maximum6, although some areas of no-analog vegetation still existed in the latter. Compared to Holocene boreal forests, cool woodlands of the late glacial contained high abundance of ash, hornbeam, spruce, and larch. Pine, alder, and birch were much rarer on the other hand.
Some argue that the rapidly changing climate of the period caused a “disequilibrium” because the peculiar mix of plant types because some plants migrate slower than others7 8. For example, imagine that the climate in a region suddenly becomes warmer. In theory, plants from further south should all migrate to claim this new real estate, but because some plants take longer to establish themselves, the initial result is a mix of warmer-climate plants living alongside cold-adapted plants that still have yet to fully vacate the area. Effectively, these were ecosystems in transition.
Another theory for why these formations existed has to do not so much with rapid changes in climate, but climate itself. According to some researchers, plants rapidly track climate changes but the climate of the late glacial period was itself so distinct from any that exist in North America that the composition of the community naturally differed6. They argue that the climate conditions of the late glacial period, with much colder winters than present, summers just as warm or warmer than present, and precipitation lower than present were sufficient to create no-analog communities, and that rapid change was not necessary. They attribute the relatively greater dissimilarity of late glacial vs. LGM pollen assemblages to modern ones to the fact that climates during the LGM have closer present-day matches (although they are now situated much further north than they were back then) than late glacial ones.
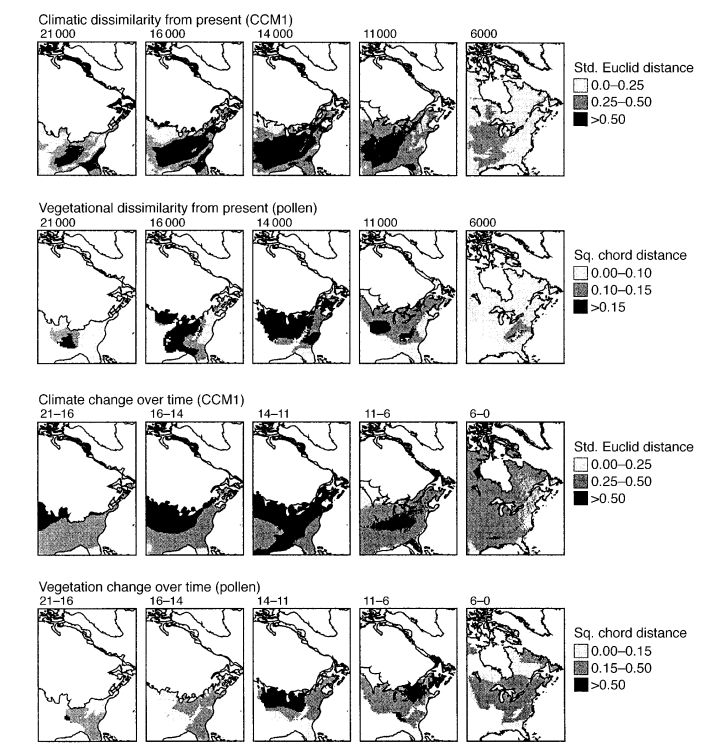
It is quite possible that it is a mix of both. I would also personally speculate that another reason for non-analog ecosystems at mid-latitudes might have been the sharp climatic gradients that existed at the time. Ice sheets extended deep into mid-latitudes during glacials, bringing very cold conditions with them, but warm water bodies were situated exactly where they are now. This meant that the distance between warm, subtropical climates near the Gulf of Mexico, and frigid, Arctic-like ones was shorter than at present. That created a sharp climatic gradient the closer one gets to the ice sheet.
The sharp gradients in temperature caused vegetation zones to become compressed latitudinally, possibly leading to a greater mixing and mingling of plants from said vegetation zones (possibly aided by stronger winds being able to carry pollen to greater distances). If this speculation has any merit, it could be that ice-sheet influenced ecosystems of the Pleistocene might have been similar to mountain ecosystems where temperatures and vegetation types change rapidly over short distances.
We also must keep in mind that extinct relatives in the forms of species or subspecies with distinct climate preferences can also create the perception of no-analog communities. For example, spruce was very widespread in eastern North America during large parts of the Pleistocene, extending far south of its present range. Someone looking at the prevalence of spruce in the southern United States may assume the climate was extremely cold. However, a major representative of the spruce in eastern North America was the now extinct Critchfield’s spruce (Picea critchfieldii), which appears to have been adapted to somewhat warmer climates than its modern relatives, which could mean that the climate was not quite as cold as it appears just from the abundance of spruce9.

Conclusion
Given the sheer size of the planet, it may seem surprising that glacial ecosystems lack close modern counterparts. But this is exactly what the paleoecological research indicates. The world during ice ages was not just a colder version of our own but a planet with quite different ecological dynamics. Most of the same species were present but often in different mixtures than at present.
Through the concept of no-analog communities-ecosystems without precise modern equivalents-we can see that species responded individually to particular climatic and environmental conditions. These assemblages are a reminder that ecological communities are not fixed entities but fluid, shifting mosaics shaped by complex interactions between climate and individual needs. No-analog conditions are not necessarily limited to glacials-some research predicts that with the planet currently experiencing a new wave of warming, we can expect unusual formations to once again emerge10. Understanding how the distributions of different plants and animals will be reshuffled will therefore be a priority for ecologists.
References
1. Wright, H. E. (1977). Quaternary Vegetation History-Some Comparisons Between Europe and America. Annual Review of Earth and Planetary Sciences, 5(1), 123–158. https://doi.org/10.1146/annurev.ea.05.050177.001011
2. Dale Guthrie, R. (2001). Origin and causes of the mammoth steppe: a story of cloud cover, woolly mammal tooth pits, buckles, and inside-out Beringia. Quaternary Science Reviews, 20(1-3), 549–574. https://doi.org/10.1016/s0277-3791(00)00099-8
3. Kahlke, R.-D. (2014). The origin of Eurasian Mammoth Faunas (Mammuthus–Coelodonta Faunal Complex). Quaternary Science Reviews, 96, 32–49. https://doi.org/10.1016/j.quascirev.2013.01.012
4. Yurtsev, B. A. (2001). The Pleistocene “Tundra-Steppe” and the productivity paradox: the landscape approach. Quaternary Science Reviews, 20(1-3), 165–174. https://doi.org/10.1016/s0277-3791(00)00125-6
5. Chytrý, M., Horsák, M., Jiří Danihelka, Nikolai Ermakov, German, D. A., Hájek, M., Hájková, P., Kočí, M., Svatava Kubešová, Pavel Lustyk, Nekola, J. C., Věra Pavelková Řičánková, Zdenka Preislerová, Philipp Resl, & Valachovič, M. (2018). A modern analogue of the Pleistocene steppe-tundra ecosystem in southern Siberia. Boreas, 48(1), 36–56. https://doi.org/10.1111/bor.12338
6. Williams, J. W., Shuman, B. N., & III, T. W. (2001). Dissimilarity Analyses of Late-Quarternary Vegetation and Climate in Eastern North America. Ecology, 82(12), 3346. https://doi.org/10.2307/2680157
7. Davis, M. B. (1986). Climatic instability, time lags, and community disequilibrium. Community ecology.
8. Knight, C. A., Blois, J. L., Blonder, B., Macias-Fauria, M., Ordonez, A., & Jens-Christian Svenning. (2019). Community Assembly and Climate Mismatch in Late Quaternary Eastern North American Pollen Assemblages. The American Naturalist, 195(2), 166–180. https://doi.org/10.1086/706340
9. Jackson, S. T., Webb, R. S., Anderson, K. H., Overpeck, J. T., Webb III, T., Williams, J. W., & Hansen, B. C. S. (2000). Vegetation and environment in Eastern North America during the Last Glacial Maximum. Quaternary Science Reviews, 19(6), 489–508. https://doi.org/10.1016/s0277-3791(99)00093-1
10. Williams, J. W., & Jackson, S. T. (2007). Novel climates, no-analog communities, and ecological surprises. Frontiers in Ecology and the Environment, 5(9), 475–482. https://doi.org/10.1890/070037