At first glance, North America and Europe share many superficial geographical and natural similarities. This is probably to be expected of any two continents, but it’s worth mentioning nonetheless. For example, both continents border the Atlantic Ocean and are located largely within the middle latitudes. The mid-Atlantic’s temperate forests somewhat resemble those of France and England, while the wet boreal forests of eastern Canada are comparable to those of northern Europe.
Southern Europe’s Mediterranean landscapes, on the other hand, are more like parts of the Pacific Coast, and Europe’s major mountain ranges-the Pyrenees, Alps, and Cantabrians-mirror those of western North America like the Rockies and Sierra Nevada. And many animal species are shared as well, such as brown bears, moose, caribou, wolves, and more, which may initially seem surprising when considering that the continents are separated by a large ocean, but not if one is aware of the legacy of the Bering Land Bridge migrations.

However, there exist differences too, even if only focusing on the mid-latitudes and ignoring the large parts of North America that have tropical or desert climates. Due to the influence of the warm Gulf Stream, locations in Europe have warmer mean annual temperatures and especially warmer winters than those at the same latitude in North America. Many people would be shocked to find out that Paris is actually slightly further north than Duluth, Minnesota, or that although Scandinavia is often stereotyped as being cold, it actually has much warmer winters than most of Canada, Alaska, or Russia. At the same time, due to being located further north, European cities aren’t as warm during summer as most North American ones.

But although there’s a lot to compare and contrast between the two continents from a natural standpoint, this is a paleo-blog and so this post will primarily focus on how these continents differed in the past, and why. Specifically, we will look at the differences between eastern North America (defined here as everything east of the Great Plains) and Europe during glacial periods, as these regions seem the most superficially similar today but were impacted by climate change in quite different ways. Then, we will take a look at what implications this had for biodiversity in both places.
So how were they different? Let’s take a look.
Grassy Europe, Wooded America
Forests are today the natural vegetative cover in Europe as well as eastern North America. In southern Europe, these are of a drier Mediterranean variety whereas central Europe and northernmost Europe contain moister temperate and boreal forests, respectively. In eastern North America, warm humid forests cover the southeastern United States while temperate and boreal forests cover central and northern latitudes, respectively.

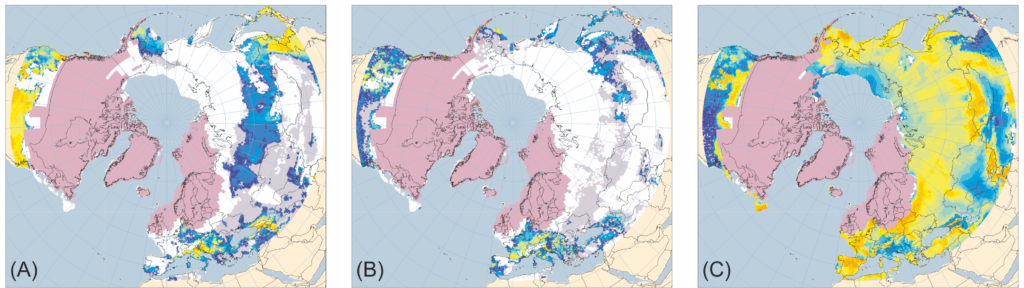
However, during the ice ages, the situation was rather different. Whereas forests became restricted in Europe, having been replaced by various open environments like steppe-tundra (especially during the coldest periods like the Last Glacial Maximum), eastern North America continued to be dominated by forests-although these were more open than at present and may be better described as parkland or woodland. Both Europe and North America contained huge ice sheets covering their northern sections, and the unglaciated regions were also far colder, so what could have caused this interesting difference?
The most important factor related to climate here is where the two regions get their precipitation from. Eastern North America largely gets its moisture from the Gulf of Mexico, whereas Europe is almost entirely dependent on the North Atlantic for its own rain and snow. This is very important as the Gulf of Mexico is a reliable and consistent source of heat and moisture, as it is a perpetually warm-water body that won’t freeze over even during glacial maxima. The result is that during glacials, evaporation continued and winds brought precipitation year-round to the eastern half of North America.

Meanwhile, the North Atlantic is very sensitive to changes in the AMOC (Atlantic Meridional Overturning Circulation), as disruptions associated with fresh-water influxes could cause the Gulf Stream to weaken or shut down. This causes the seas west of Europe to become colder and has downwind effects on the continent. The difference would be especially noticeable in winter as an active Gulf Stream prevents Europe from cooling down as much as would be expected for its latitude. This is what happened in the millennial-scale cold pulses of the ice-ages known as “stadials”.
Colder sea surface temperatures had the effect of reducing evaporation and hence precipitation. Extensive sea ice, rather than open water, dominated the North Atlantic Ocean in winter which meant no moderating oceanic influence on temperature and almost no evaporation. This led to a disproportionate drop in temperature and precipitation for Europe during that season. Hence, western and central Europe-which today have relatively modest differences between summer and winter temperatures and evenly spread out rainfall-had stadial climates that were drier, colder, and more seasonal like modern Siberia or Central Asia.
The result is that while eastern North America received enough precipitation and warmth to support forests, in Europe the harsher conditions caused them to diminish. This pattern largely does not apply to Europe during interstadial periods when the AMOC and hence Gulf Stream was strong, but most interstadials were probably not long enough for forests to establish themselves before the next cold snap (stadial) killed them. This created or at least contributed to the conditions for a general contraction of forest cover across much of the continent for tens of thousands of years during the last glacial.
Still, there is some nuance here with regard to the exact nature and causes of glacial vegetation in Europe that we will discuss next before expanding on the comparisons between that continent and North America.
A (Somewhat) Unclear Picture?
The Last Glacial Maximum around 21 thousand years ago was the coldest part of the last ice age and while there is broad agreement that open environments dominated across most of Europe during this period, there is considerable disagreement regarding the extent of woodland and forest refugia north of the Alps. The Carpathians are often claimed to be an area that may have contained favorable microclimates for forests. One study from 2024 by Hošek et al. argues even temperate trees like Quercus (Oak), Alnus (Alder), and Ulmus (Elm) survived near isolated geothermal hot springs in central Europe. But regardless of whether forest refugia persisted in certain locations, vast swathes of unglaciated Europe were ultimately still dominated by cold steppe-like environments during the LGM.
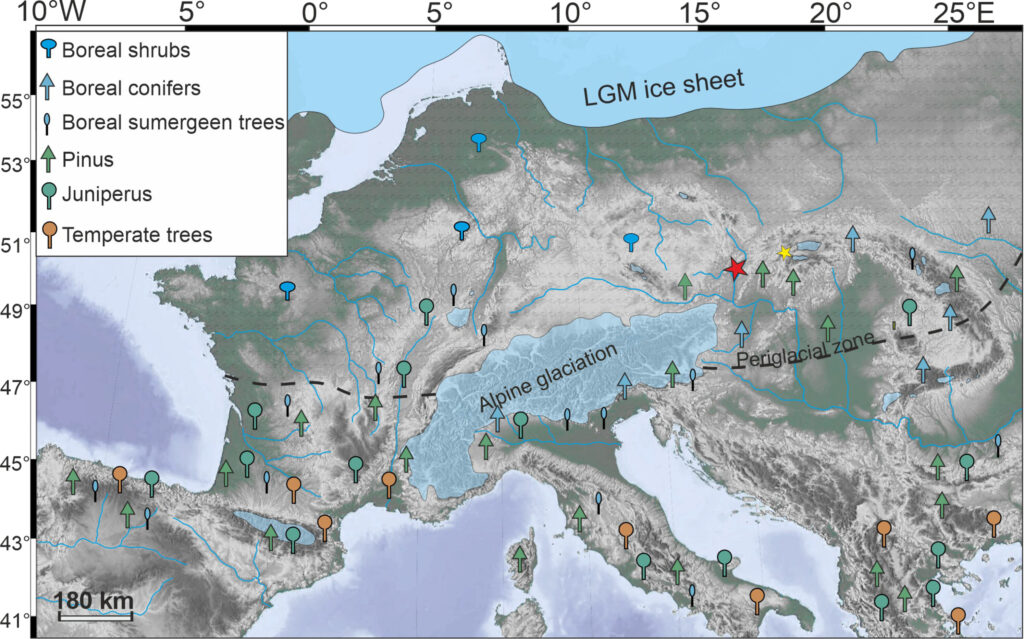
The causes for the prevalence of open steppes are another cause of contention. Summer temperatures were definitely a limiting factor, as trees need the warmest month of the year to be at least 10°C (50°F) average to survive. Summers in European locales are not as warm as in North American ones with the same mean annual temperature, so comparable decreases in summer temperature could shift the tree-line a greater distance in Europe than in North America. The tree-line being further south in western Europe than eastern Europe at the time, the latter being more continental with warmer summers, further reflects this trend. Exceptionally low CO2 values during the LGM (185 ppm) may have also contributed to openness by reducing the water-use efficiency of plants and hence favoring grasses and forbs over trees and shrubs.
But one intriguing study (Kaplan et al., 2016) argues that natural factors like climate, CO2 concentration, and disturbance by megaherbivores can’t fully explain the contraction of forest in Europe at this time on their own and posits that human-induced fire was also necessary. At the moment, I am not fully convinced of this hypothesis simply because a large part of Europe was seemingly abandoned by people during the LGM, especially north of the Alps (the Gravettian culture for example disappeared).
Still, it’s a fascinating thought and all of this shows how difficult it can be not just to precisely determine past environmental conditions but also to disentangle the precise causes of said conditions. I strongly suspect that interactive effects between multiple forces were decisive in promoting the open character of vegetation in glacial Europe. Vegetation models that explicitly account for this would come in handy.
Anyway, back to the comparisons. Unlike Europe, unglaciated eastern North America-despite also being very cold compared to today-was dominated by woodland and forest biomes even during the LGM. The map below shows how close to the ice sheet margin that boreal forest and forest tundra (transitional between boreal forest and tundra) grew in eastern North America ~21-22 thousand years ago, which indicates that the supply of moisture and summer warmth was plentiful relative to Europe. Note that although the map only shows northern North America, pollen data indicates woody biomes extended all the way down to the coast of the Gulf of Mexico.
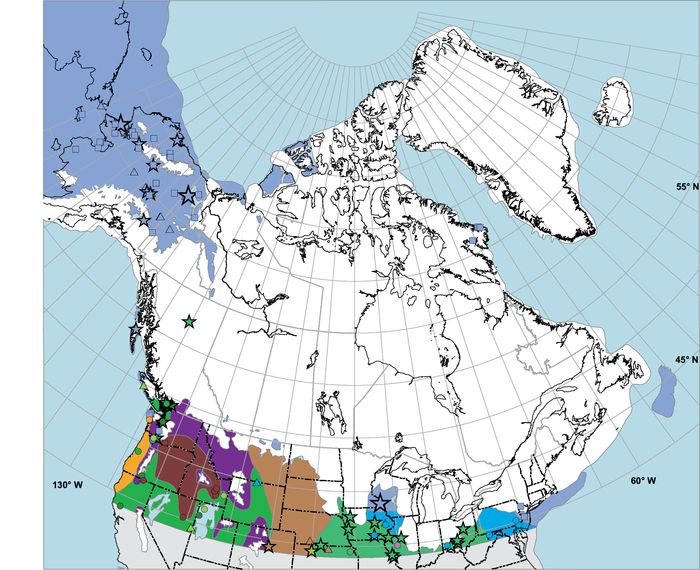
Tree cover was, however, sparser during the last glacial in North America than during the Holocene and this may have been due to climate, lower CO2, the presence of large herbivore populations, or some mix of these factors. Regardless, tree populations did well throughout the Pleistocene in comparison to their European counterparts, which experienced gradual losses in diversity of taxa.
Barriers and Consequences
Under quintessential glacial conditions, Europe was generally not a hospitable place for trees as the combination of cold and reduced moisture availability made vast swathes of the continent harsh for them. This would have been especially true for certain types of thermophilous (warmth-loving) trees which would have needed to disperse to climatically favorable refugia, such as in southern Europe.
But shifting their ranges to southern Europe is not exactly easy. For one, the paths from central Europe to the Mediterranean peninsulas (Iberia, Italy, and the Balkans) are bounded by towering east-west mountain ranges like the Pyrenees, the Alps, and the Carpathians, and the peninsulas themselves are quite mountainous with numerous ranges such as the Apennines, Dinaric Alps, and Pindus which further complicate migration to the coastal plains. Anatolia, connected to the Balkans by land at the time, is also highly mountainous.

Moreover, southern Europe has different rainfall patterns than northern Europe, with dry summers and wet winters rather than consistent year-round moisture. Even worse, millennial-scale stadial events, especially Heinrich stadials, brought drier conditions not only to northern Europe but southern Europe as well, as indicated by the contraction of tree taxa and expansion of steppe from Spain to Greece during these intervals. This meant that thermophilous trees that made it to southern Europe would have had the additional problem of needing to survive water-stress.
The double whammy of geographic barriers to migration and turbulent climate change caused Europe to lose a larger share of tree types since the start of major northern hemisphere glaciations a few million years ago than did eastern North America or East Asia. Some arboreal genera that went extinct in Europe but survived in North America and/or East Asia include Carya (Hickory), Engelhardia, Tsuga (part of the pine family), Taxodium (part of the Cypress family), Chamaecyparis (false cypress), and Liriodendron (the tulip tree). Others that were once common in Europe but have since become very restricted include Liquidambar (sweetgum), Zelkova, and Pterocarya (wingnut), all of which are now only natively found in the fringes of Europe such as islands, the southernmost parts of peninsulas, or the Caucasus.

The eastern United States had practically no meaningful barriers between the warm coast of the Gulf of Mexico in the south and the Laurentide Ice Sheet to the north. The land in eastern North America to the east, west, and south of the Appalachian mountains is overwhelmingly flat. The Appalachians themselves are not very tall and run north to south rather than east to west, so they are not a real hurdle. The Gulf of Mexico, as we mentioned, remained a reliable source of moisture and humid subtropical climates persisted, just further south than at present. This gave flora and fauna an easy way to survive cooling (or warming) via migration.
For another geographical contrast, East Asia was also influenced by important temperature and moisture changes during the Quaternary-most importantly with regard to the East Asian monsoon which periodically weakened and shifted further south. However, the northern, central, and southern parts of East Asia are relatively well-connected to each other. Mainland East Asia is a large landmass which-for the most part-lacks the the same kinds of narrow chokepoints leading into peninsulas found in Europe, and the mountains in eastern China are not as imposing as the Alps or Pyrenees. This, along with the absence of major ice sheets in East Asia (aside from very high-elevations) during glaciations, explains the high degree of plant richness there today.
Based on what I’ve written so far in this post, it may appear that mountains are inherently a hurdle to maintaining biodiversity, but this is not always true. In fact, it can often be the opposite. While mountainous regions can be barriers to migration, they also host a variety microclimates due to topographical complexity, allowing refugia for various plant types to flourish. Possible forest microrefugia in the Carpathians have been referenced here earlier, and it’s thought that the high tree diversity in the Balkans is a function of that region’s geographical heterogeneity with high mountains and low valleys. Another important factor was the limited or absent glaciations in the Balkan mountains compared to the Alps or Pyrenees. The mountains of southwestern China have exceptional tree and overall plant diversity for similar reasons.
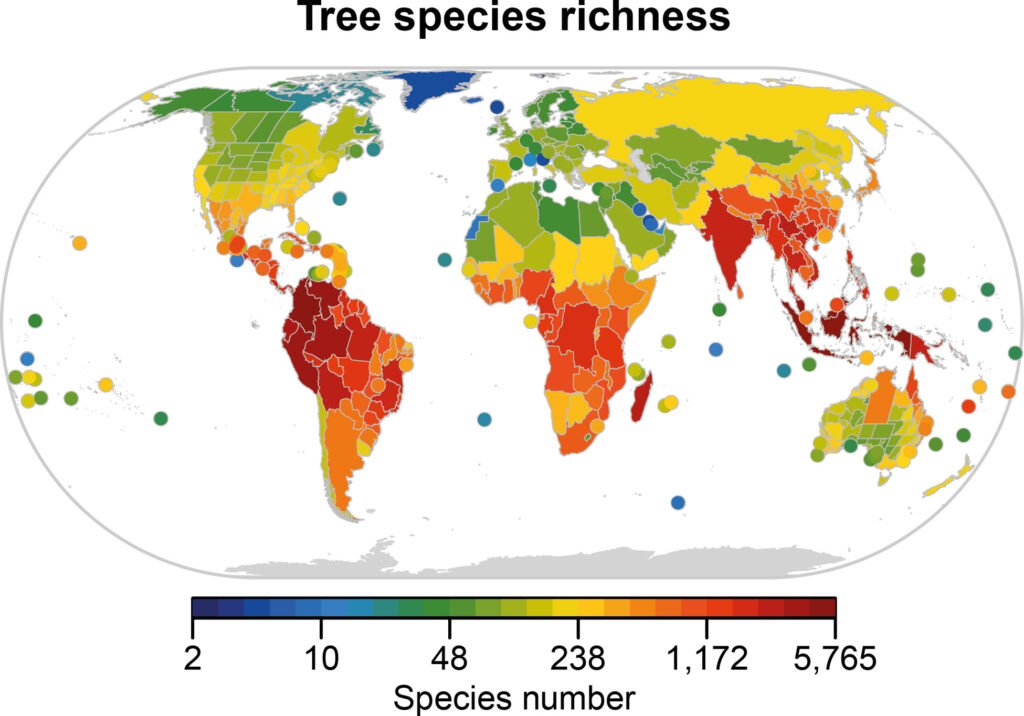
Regardless, the (left) map below shows that tree species richness in Europe is lower than in eastern North America and much lower than in East Asia. Keep in mind that this does not apply to plants as a whole, as Europe does contain decent plant diversity (right map below), which could be because trees are more vulnerable to climatic instability than other types of vegetation. This would make sense given that trees as large and long-living organisms would generally have stricter requirements for survival than smaller plants with shorter lifespans.
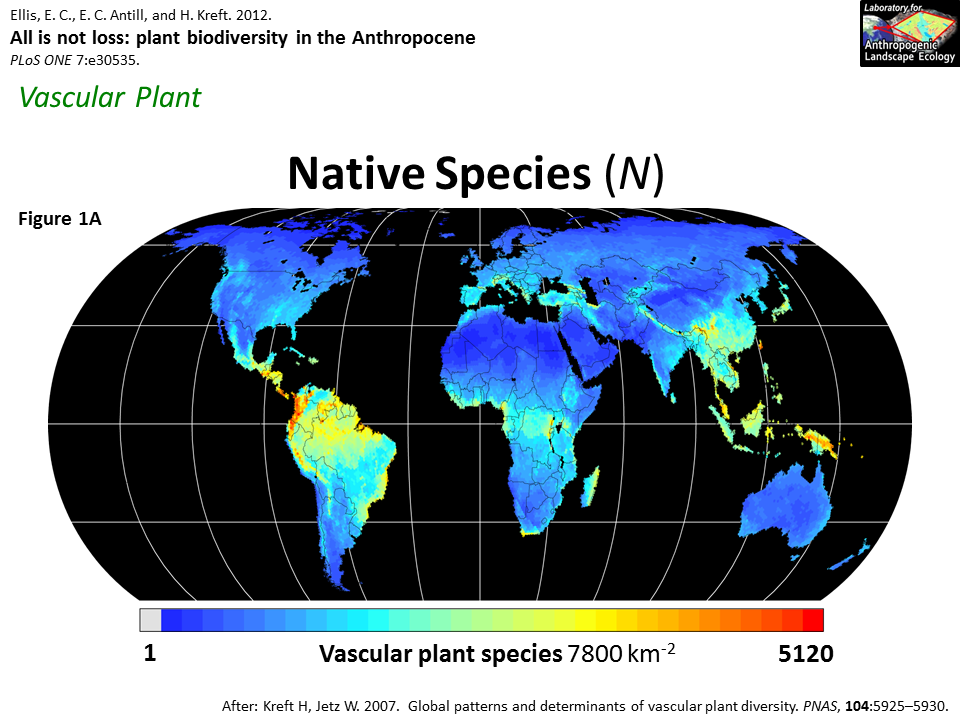
The legacy of these geographical differences on dispersal has been observed at the genetic level as well. Tree species in northern Europe have less genetic diversity than their counterparts in southern Europe, whereas no latitudinal gradient in genetic diversity exists for eastern North America. The distinction between the two continents is probably caused by southern Europe being the refuge for many tree species, which diverged in isolation from each other, and then experienced bottlenecks as they migrated northwards across various geographic barriers. Meanwhile, tree populations (even temperate species) in eastern North America persisted in northern areas not far from the ice sheet through the coldest periods, and the lack of barriers prevented bottlenecks during plant dispersal from southern refugia to the north.
So it’s clear that Europe was hard mode for trees trying to survive Quaternary climate fluctuations. But what about animals? Animals of course experienced many parallel large-scale range contractions, as it’s well known that temperate animals migrated to southern refugia during cold periods and recolonized northern areas during interglacials. But there also seems to have been a trend in some extinct animals of downgrading in size as Pleistocene glacial cycles intensified over time. Both Hippopotamus antiquus and Panthera spelaea (cave lions) are suggested to have become smaller through the Middle Pleistocene due to repeated and increasingly severe climatic fluctuations reducing consistent food availability.
If this trend is genuine rather than a sampling artifact and is reflected in other European animal taxa as well, it would be yet another reflection of how interconnected ecosystems are, with stresses on plant communities mirroring stresses on animal communities in a shockingly strong way.
Conclusion
Although there are many commonalities between Europe and eastern North America from a natural viewpoint, the way the two regions experienced past climatic changes was very different. Geographical realities created a scenario where their paleoenvironments were rather distinct from each other during the Pleistocene.
Wooded habitats remained extensive in eastern North America at all periods whereas they contracted severely in Europe during ice ages. Moreover, the pathways for biotic migration in response to climate change in eastern North America, like in East Asia, were far superior to those in Europe which had extensive geographical barriers and ended up retaining less of its biodiversity as a result.
This is one of my favorite examples of how geography has shaped the natural world over time in somewhat surprising ways.
References
Basil, Fasel, M., Kaplan, J. O., Russo, E., & Burke, A. (2024). The climate and vegetation of Europe, northern Africa, and the Middle East during the Last Glacial Maximum (21 000 yr BP) based on pollen data. Climate of the Past, 20(9), 1939–1988. https://doi.org/10.5194/cp-20-1939-2024
Binney, H., Edwards, M., Macias-Fauria, M., Lozhkin, A., Anderson, P., Kaplan, J. O., Andreev, A., Bezrukova, E., Blyakharchuk, T., Jankovska, V., Khazina, I., Krivonogov, S., Kremenetski, K., Nield, J., Novenko, E., Ryabogina, N., Solovieva, N., Willis, K., & Zernitskaya, V. (2017). Vegetation of Eurasia from the last glacial maximum to present: Key biogeographic patterns. Quaternary Science Reviews, 157, 80–97. https://doi.org/10.1016/j.quascirev.2016.11.022
Denton, G., Alley, R., Comer, G., & Broecker, W. (2005). The role of seasonality in abrupt climate change. Quaternary Science Reviews, 24(10-11), 1159–1182. https://doi.org/10.1016/j.quascirev.2004.12.002
Duprat-Oualid, F., Bégeot, C., Peyron, O., Rius, D., Millet, L., & Magny, M. (2022). High-frequency vegetation and climatic changes during the Lateglacial inferred from the Lapsou pollen record (Cantal, southern Massif Central, France). Quaternary International, 636, 69–80. https://doi.org/10.1016/j.quaint.2022.04.012
Dyke, A. S. (2007). Late Quaternary Vegetation History of Northern North America Based on Pollen, Macrofossil, and Faunal Remains*. Paleoenvironments, 59(2-3), 211–262. https://doi.org/10.7202/014755ar
Ellis, E. C., Antill, E. C., & Kreft, H. (2012). All Is Not Loss: Plant Biodiversity in the Anthropocene. PLoS ONE, 7(1), e30535. https://doi.org/10.1371/journal.pone.0030535
Fastovich, D., Radeloff, V. C., Zuckerberg, B., & Williams, J. W. (2024). Legacies of millennial-scale climate oscillations in contemporary biodiversity in eastern North America. Philosophical Transactions of the Royal Society B Biological Sciences, 379(1902). https://doi.org/10.1098/rstb.2023.0012
Gábor Újvári, Bernasconi, S. M., Stevens, T., Sándor Kele, Barna Páll-Gergely, Gergely Surányi, & Attila Demény. (2021). Stadial‐Interstadial Temperature and Aridity Variations in East Central Europe Preceding the Last Glacial Maximum. Paleoceanography and Paleoclimatology, 36(8). https://doi.org/10.1029/2020pa004170
Gömöry, D., Zhelev, P., & Brus, R. (2020). The Balkans: a genetic hotspot but not a universal colonization source for trees. Plant Systematics and Evolution, 306(1). https://doi.org/10.1007/s00606-020-01647-x
Guthrie, D., & van Kolfschoten, T. (2000). Neither warm and moist, nor cold and arid: the ecology of the Mid Upper Palaeolithic. Hunters of the golden age: the mid upper palaeolithic of Eurasia, 30, e20.
Hošek, J., Pokorný, P., Storch, D., Kvaček, J., Havig, J., Novák, J., Hájková, P., Jamrichová, E., Brengman, L., Radoměřský, T., Křížek, M., Magna, T., Rapprich, V., Laufek, F., Hamilton, T., Pack, A., Di Rocco, T., & Horáček, I. (2024). Hot spring oases in the periglacial desert as the Last Glacial Maximum refugia for temperate trees in Central Europe. Science Advances, 10(22). https://doi.org/10.1126/sciadv.ado6611
Huntley, B., Allen, J. R. M., Collingham, Y. C., Hickler, T., Lister, A. M., Singarayer, J., Stuart, A. J., Sykes, M. T., & Valdes, P. J. (2013). Millennial Climatic Fluctuations Are Key to the Structure of Last Glacial Ecosystems. PLoS ONE, 8(4), e61963. https://doi.org/10.1371/journal.pone.0061963
Ilvessalo, L. (1927). Cultivation of foreign species of trees. Silva Fennica, 4. https://doi.org/10.14214/sf.a8390
Jackson, S. T., Webb, R. S., Anderson, K. H., Overpeck, J. T., Webb III, T., Williams, J. W., & Hansen, B. C. S. (2000). Vegetation and environment in Eastern North America during the Last Glacial Maximum. Quaternary Science Reviews, 19(6), 489–508. https://doi.org/10.1016/s0277-3791(99)00093-1
Juřičková, L., Horáčková, J., Horsáková, V., Hájková, P., Hošek, J., & Horsák, M. (2025). Persistence and dynamic of forest snails in the Western Carpathians over the last 40 thousand years. Boreas, 55(1), 98–109. https://doi.org/10.1111/bor.70019
Kaplan, J. O., Pfeiffer, M., Kolen, J. C. A., & Davis, B. A. S. (2016). Large Scale Anthropogenic Reduction of Forest Cover in Last Glacial Maximum Europe. PLOS ONE, 11(11), e0166726. https://doi.org/10.1371/journal.pone.0166726
Keppel, G., Craven, D., Weigelt, P., Smith, S. A., van der Sande, M. T., Sandel, B., Levin, S. C., Kreft, H., & Knight, T. M. (2021). Synthesizing tree biodiversity data to understand global patterns and processes of vegetation. Journal of Vegetation Science, 32(3). https://doi.org/10.1111/jvs.13021
Lumibao, C. Y., Hoban, S. M., & McLachlan, J. (2017). Ice ages leave genetic diversity “hotspots” in Europe but not in Eastern North America. Ecology Letters, 20(11), 1459–1468. https://doi.org/10.1111/ele.12853
Magri, D., Di Rita, F., Aranbarri, J., Fletcher, W., & González-Sampériz, P. (2017). Quaternary disappearance of tree taxa from Southern Europe: Timing and trends. Quaternary Science Reviews, 163, 23–55. https://doi.org/10.1016/j.quascirev.2017.02.014
Marciszak, A., Ivanoff, D. V., Semenov, Y. A., Talamo, S., Bogdan Ridush, Veiber, A., Yanish, Y., & Kovalchuk, O. (2022). The Quaternary lions of Ukraine and a trend of decreasing size in Panthera spelaea. Journal of Mammalian Evolution, 30(1), 109–135. https://doi.org/10.1007/s10914-022-09635-3
Mazza, P. P. A., & Bertini, A. (2012). Were Pleistocene hippopotamuses exposed to climate-driven body size changes? Boreas, 42(1), 194–209. https://doi.org/10.1111/j.1502-3885.2012.00285.x
McLachlan, J. S., Clark, J. S., & Manos, P. S. (2005). Molecular Indicators of Tree Migration Capacity Under Rapid Climate Change. Ecology, 86(8), 2088–2098. https://doi.org/10.1890/04-1036
Prentice, I. C., & Harrison, S. P. (2009). Ecosystem effects of CO2 concentration: evidence from past climates. Climate of the Past, 5(3), 297–307. https://doi.org/10.5194/cp-5-297-2009
Tallavaara, M., Luoto, M., Korhonen, N., Järvinen, H., & Seppä, H. (2015). Human population dynamics in Europe over the Last Glacial Maximum. Proceedings of the National Academy of Sciences, 112(27), 8232–8237. https://doi.org/10.1073/pnas.1503784112
Tzedakis, P. C., Emerson, B. C., & Hewitt, G. M. (2013). Cryptic or mystic? Glacial tree refugia in northern Europe. Trends in Ecology & Evolution, 28(12), 696–704. https://doi.org/10.1016/j.tree.2013.09.001
Tzedakis, P. C., Frogley, M. R., Lawson, I. T., Preece, R. C., Cacho, I., & de Abreu, L. (2004). Ecological thresholds and patterns of millennial-scale climate variability: The response of vegetation in Greece during the last glacial period. Geology, 32(2), 109. https://doi.org/10.1130/g20118.1
Tzedakis, P. C., McManus, J. F., Hooghiemstra, H., Oppo, D. W., & Wijmstra, T. A. (2003). Comparison of changes in vegetation in northeast Greece with records of climate variability on orbital and suborbital frequencies over the last 450 000 years. Earth and Planetary Science Letters, 212(1-2), 197–212. https://doi.org/10.1016/s0012-821x(03)00233-4
Williams, J. W. (2003). Variations in tree cover in North America since the last glacial maximum. Global and Planetary Change, 35(1-2), 1–23. https://doi.org/10.1016/s0921-8181(02)00088-7
Williams, J. W., Shuman, B. N., & III, T. W. (2001). Dissimilarity Analyses of Late-Quarternary Vegetation and Climate in Eastern North America. Ecology, 82(12), 3346. https://doi.org/10.2307/2680157
Wren, C. D., & Burke, A. (2019). Habitat suitability and the genetic structure of human populations during the Last Glacial Maximum (LGM) in Western Europe. PLOS ONE, 14(6), e0217996. https://doi.org/10.1371/journal.pone.0217996