During glacial periods when the global sea level was lower, large areas of now-submerged land were exposed. In many cases, the exposure of these continental shelves allowed landmasses that are now disconnected by water to become joined with one another. One of the most famous cases of this was the paleocontinent “Sahul”, which connected the mainland of Australia with the islands of Tasmania and New Guinea.
For a reader familiar with the term, the word “Sahul” immediately conjures a sense of primordiality, and this is not without justification. Humans arrived there tens of thousands of years earlier than they did in the Americas, and remained almost entirely cut off from the rest of the world until only a few centuries ago. With some exceptions, hunting-gathering remained the dominant mode of life in the region for much longer than in other continents. The paleocontinent included a great variety of habitats from the dense rainforests of New Guinea to the bone-dry deserts of central Australia. Many of these environments were challenging to survive in, meaning that they could support only sparse human populations.
I’ve written about Australia before but the broader region is fascinating enough that it warrants another article dedicated to it. In this post, I will analyze controversies surrounding the initial settlement of Sahul, which I’ve already partly touched on before. Subjects will include determining the timing of human settlement, what we know about Australasian demographic history, from admixture with Denisovans to claims of an Indian connection, the story of dingoes, how fire shaped Australian landscapes, and a bit more.

Let’s start from the beginning of the human story in Sahul.
Breaching the Weber Line and Ancestral Origins
Reaching Sahul through land alone was not possible even during the sea level lowstands of the Pleistocene, because deep water boundaries separated Sundaland (the connected landmass of Southeast Asia) and Sahul. In fact, these water barriers have resulted in minimal faunal and floral exchange between the Asian and Australasian biogeographic realms. The “Wallace Line” represents the line between Asia and Wallacea. Wallacea is a region which includes the islands of Sulawesi, Sumba, Lombok, Timor, and Flores that is florally and faunally transitional between the Indomalayan and Australasian realms. Meanwhile, the hard boundary that forms the western limit of Australasia in the strict sense is represented by either the Weber or Lydekker line.
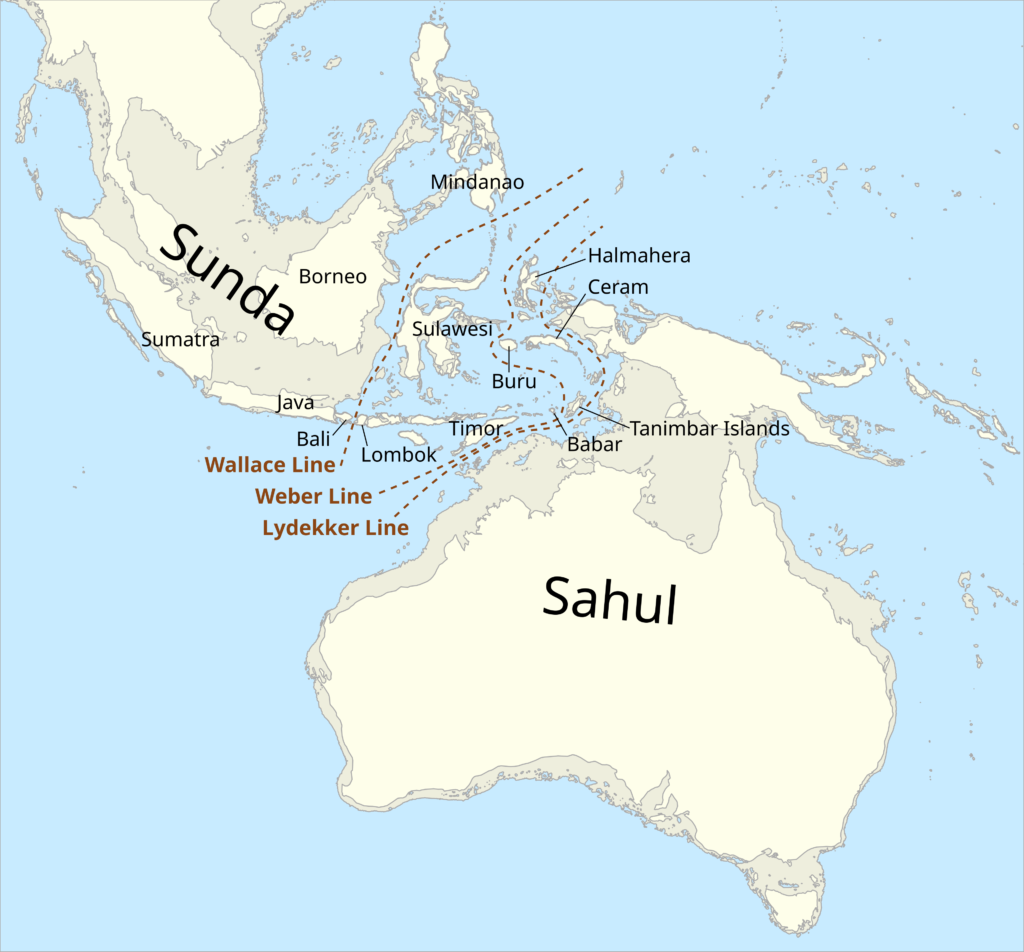
Many creatures of Afro-Eurasian origin made it past the Wallace line and into islands like Sulawesi, Flores, and Timor such as suids (like the Babirusa), proboscideans (stegodons), and hominins (in the form of Homo floresiensis and perhaps other species). But crossing the Weber and Lydekker lines proved far more challenging. Among the few species that are confirmed to have penetrated this barrier and made it into Sahul-New Guinea, Australia, and Tasmania-is our own kind: Homo sapiens.
In the world of archeology, there is controversy over when humans arrived in any given region or continent. Sahul is obviously no exception, and in fact the debate surrounding the timing of settlement in this case is one of the most heated. There are two hypotheses in this regard: one is that people first arrived in Sahul at least 65 thousand years ago and the other is that humans arrived shortly after 50 thousand years ago. These competing arrival dates are both quite ancient and would indicate that at least some human populations had developed the ability to make watercraft capable of long-distance travel across deep water rather early on. But which of them is actually correct?
Madjedbebe: Not So Fast
The claim of human arrival at 65 kya or earlier initially rested solely on one site, the Madjedbebe rock shelter in Arnhem Land, northern Australia 1. The reported date produced a wave of excitement in the press and among archeological circles, and it has been frequently cited since. Soon after, some sites in Southeast Asia-on the path to Sahul-were also claimed to be roughly as old 2 3, potentially offering some support to the notion of such an early settlement of Sahul.

The 65 kya date for the Madjedbebe site has been strongly criticized by prominent archeologists, as have the ones in Southeast Asia that are claimed to be roughly contemporaneous 4 5. Moreover, such an early settlement of Sahul is at odds with what we know about the most recent Out of Africa migration and most of the genetic evidence on Australo-Papuans, which instead argues for a later settlement 6 7 8. The Initial Upper Paleolithic occurred at around 50 thousand years ago, and coincides with when humans began spreading across Eurasia. As humans expanding from Africa must inevitably cross Eurasia to reach Australia, it stands to reason that human settlement of the latter could not have occurred earlier than this.
But this assumption only holds true if there weren’t prior Out of Africa (OoA) events involving anatomically modern humans that reached far, which is why supporters of a 65 thousand year or earlier settlement argue for a separate dispersal which occurred prior to the main OoA wave that produced Eurasians. This proposed early migration would have followed the coasts of Asia, from Arabia through India and Indochina before finally reaching Sahul and giving rise to groups that were either fully or partly the ancestors of Sahulian populations.
To be fair, some genetic research years ago claimed that the ancestors of Australian Aboriginals and Papuans diverged from other non-Africans prior to the split between East Asians and Europeans 9. That could point to those groups being the result of an earlier Out of Africa expansion. If true, this gives an opportunity to justify the 65 thousand year old settlement age and the hypothesized early dispersal, along with claims that Australian Aboriginals represent the oldest continuous human population outside of Africa.
However, the problem is that the relatively large amount of Denisovan ancestry found in Australian and Papuan genomes was not accounted for. As Denisovans split off from Homo sapiens hundreds of thousands of years ago, elevated Denisovan ancestry in populations could cause those populations to seem as though they diverged earlier than they actually did, explaining why Australasian groups appeared to be an outgroup to both East Asians and Europeans.
Later analyses, which controlled for the Denisovan input, actually found Australasian populations to be a sister-group to East Asians 7 10 11. One 2016 study used a method called Relative Cross Coalescent Rate to argue that Papuans were at least partly derived from an earlier population 12, but this too was later refuted when a subsequent study indicated that was an illusion created by a strong bottleneck and slow population growth among Papuans 7. After accounting for Denisovan admixture and population specifics, Oceanian genetics can be explained without the need to invoke an earlier OoA contribution, although a genuine but negligible input from one is still a possibility.
Furthermore, analyses of Neanderthal ancestry in modern humans, including Papuans and Australians, indicate that all non-Africans have approximately 2% Neanderthal ancestry which appears to be from the same admixture event roughly 50 thousand years ago 11 13. It would be strange if Sahulian populations descended wholly or substantially from an earlier wave of migration but managed to retain exactly the same percentage of Neanderthal ancestry as other non-Africans, with the date of admixture being exactly the same.
Therefore, we can say that Australian Aboriginal and Papuan people do indeed descend from the same wave of Out-of-Africa expansion that gave rise to all other non-Africans. If there were humans in Australasia (or Southeast Asia) long before 50 kya, these are likely to represent old lineages that died off or managed to only leave tiny, probably undetectable traces of ancestry in modern populations.
Denisovan Admixture
Denisovans are a sister lineage to Neanderthals, named after the Denisova Cave in southern Siberia. The two split off from each other after their common ancestor split from the lineage leading to Homo sapiens 14. Whereas Neanderthals occupied large parts of western Eurasia, Denisovans occupied much of the east, and interbreeding seems to have occurred in areas where they overlapped 15. Both also mixed with Homo sapiens after the latter left Africa, but while non-Africans all contain similar amounts of Neanderthal ancestry, Denisovan ancestry is more variable.
It’s well known that this ancestry peaks in Oceanian populations- it comprises 3-6% of their genome in contrast to the less than 1% found in mainland East Asians 16. Denisovans had a broad range and it’s been shown that the Denisovan ancestry found in Oceanians diverged long ago from the original type specimen from Denisova Cave in southern Siberia 14. This only makes sense; if Oceanians picked up substantial Denisovan ancestry, it was probably somewhere proximate to Sahul, such as Sundaland, where we’d expect the local Denisovans to be distinct from those living much further north 16 17.
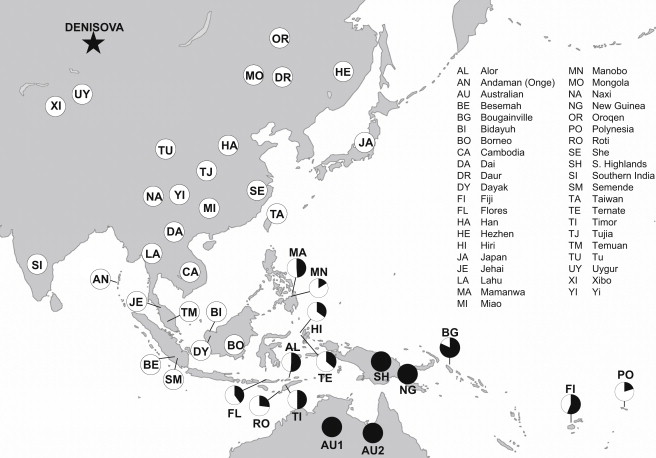
What is most perplexing, perhaps, is the timing of Denisovan admixture. Some put the date for the introgression close to the settlement time of Sahul-no later than 44 thousand years ago 18, which sounds reasonable. But other studies have looked at Papuan genomes and argued for Denisovan introgression occurring as late as 30 thousand years ago, with possibly multiple introgression events involving Denisovans 7 19. I personally find the latter suggestion very odd and struggle to come up with clear explanations for how it would have occurred.

This is a rather recent date assuming Papuans did in fact settle New Guinea shortly after 50 thousand years ago, because this would mean almost 20 thousand years passed before the final pulse of mixing and that it occurred within New Guinea rather than in Southeast Asia. New Guinea, being past the Weber/Lydekker line, would have been difficult for archaic hominins to reach, and such a late date raises questions about why it took so long for the mixing to occur. Papuans are the main focus when it comes to Denisovan ancestry in Oceanian populations, but I think analyzing and comparing the Denisovan ancestry in Australian Aboriginals would offer more context. Precisely refining the timing of Denisovan admixture is a priority.
Regardless, following settlement, there appears to be a trend of slowly increasing population over time in Australian populations 6, but the same does not appear to be true for Papuans who experienced a strong bottleneck and slow population growth 7. The difference in population growth could simply be due to the larger size of Australia compared to New Guinea and/or the challenges of surviving in the rainforest environments of the latter. Additionally, all groups in Sahul display a strong degree of regionalism genetically based on mitogenomes 6 20, meaning there was minimal mixing between people in different geographical locations.
Indian Ancestry
Misconceptions abound regarding the demographic history of Sahul. In addition to the previously mentioned dubious idea that the native populations descend from an earlier OoA migration, one commonly held belief involves Australian Aboriginals specifically. In many online circles, it is often claimed that a strong ancestral connection of one form or another exists between Australian Aboriginals and Indians.
19th century European anthropologists, most prominently TX Huxley, noted the physical similarity between indigenous Australians and the people of South India 21. The alleged morphological similarities pushed a few geneticists to investigate potential links between the two groups. One study claimed there were similarities in Y-DNA 22 and another used whole-genome analysis to claim that Australian Aboriginals carry ancestry from a wave of Indian migration, most likely from South India, to Australia that occurred during the Holocene some 4,000 years ago or so, around the time of the arrival of the dingo 23. The findings of the study were reported in the news.
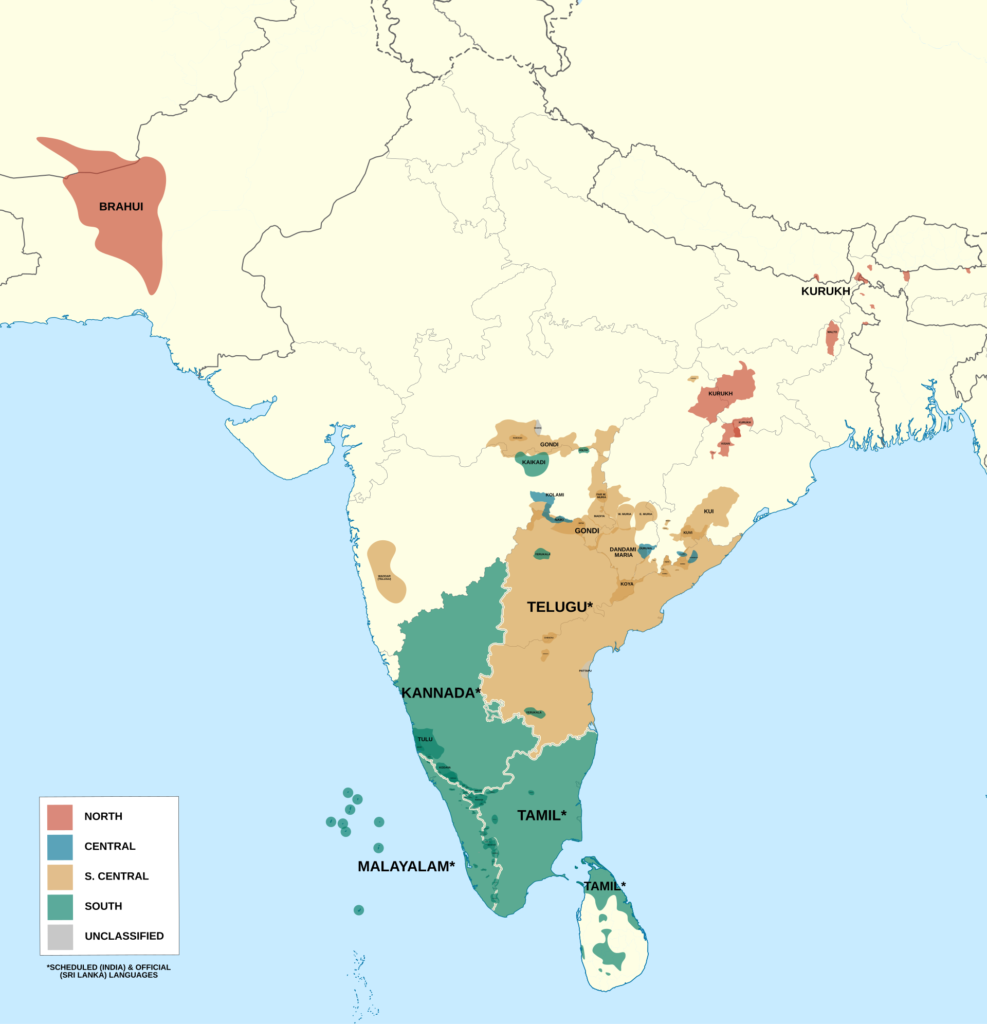
As fascinating as some sort of special connection between native Australians and Indians would be, this sadly appears to be an outdated myth. Subsequent studies on paternal and maternal haplogroups have failed to reveal any such admixture during the Holocene, and instead argue for a long-term isolation of Papuan and Australian people since the time of settlement shortly after 50 thousand years ago 24 25 26 27. As for the alleged phenotypic similarities? We can chalk them up to being the result of convergent selection or genetic drift, something that was not accounted for by early anthropologists who were working within a cruder framework for understanding relations between different human populations.
It’s a bit disappointing that it turns out that Indians don’t have long-lost family in the land down under. But that’s not to say that there’s no connection at all. Actually, part of the ancestry of Indians originates from a lineage that is distantly linked to East Asians, Andaman Islanders, and Oceanians. This lineage that Indians partly descend from is part of the East Eurasian branch, as are East Asians, Andaman Islanders, and Oceanians, as opposed to West Eurasian (i.e. Europeans and West Asians) 28. The two branches split off sometime after the Out of Africa expansion. Hence, Oceanians are slightly closer to Indians and East Asians than they are to Europeans, but the connection doesn’t go much deeper than that.
The exact timing of Sahul’s settlement and genetic history of Australasian populations will continue to be debated. A picture, however vague, does seem to be emerging in the overall literature. But the indigenous human settlers of Sahul aren’t the only arrivals whose precise origins have been the subject of controversy: the dingo, a representative of the dog lineage (man’s best friend), is another.
Dingo Invasion

Unlike wolves and coyotes, dingoes have a physical appearance that-to the untrained eye-would easily allow them to pass as a regular breed of domestic dog. But unlike domestic dogs, dingoes live in the wild and they do not bark. Yes, dingoes don’t bark-they howl.
Dingoes arrived in Australia much later than humans. It’s estimated that they have been in Australia for about 3,500 years based on radiocarbon dating of the oldest dingo remains on the continent 29. This would make them surprisingly late arrivals to the continent. Although dingoes are wild, they fall within the domestic dog genetic clade and the inherent difficulty of reaching Australia means that humans must have brought them there at some point, in the form of dogs that were once domesticated 30 31. This means that some exchange, however minor, did occur between Australia and the rest of the world prior to the colonial era.
Who actually brought these odd canines there has been a mystery. Some speculated, based on phenotypic similarities with Indian pariah dogs, that dingo ancestors may have arrived from India 32. That aligns with the outdated speculation of some sort of human migration from that part of the world. But just like the claims of Indian ancestry in native Australians, the notion of Indian dogs being the progenitors of dingoes doesn’t hold up.
Genetic testing shows dingoes to be closest to New Guinea singing dogs, and both dingoes and singing dogs are closest to domestic dogs from Southeast Asia and southern China 30 31. Hence, the migration that gave rise to dingoes and singing dogs occurred from somewhere in East or Southeast Asia rather than South Asia, which accords with geographical proximity. It’s worth noting that neither singing dogs nor dingoes bark, unlike Indian pariah dogs who do, again casting doubt on the latter being the progenitor of either of the former two. Even then, it’s unclear which human group exactly was responsible for bringing the dogs from East/Southeast Asia into New Guinea and Australia.
What effects did the dingo’s entry into Australia have on the local fauna? The thylacine, also known as the Tasmanian tiger, went extinct from Tasmania in the early 20th century due to persecution by humans. Despite being called a tiger and having a superficial resemblance to dogs, it was neither, and was actually a native marsupial predator. It was once extant not only in Tasmania but also mainland Australia and New Guinea but went extinct in the latter places thousands of years ago.

The disappearance of not just the thylacine but also the Tasmanian devil and Tasmanian hen from mainland Australia coincided with the dingo’s entry into Australia. Tasmanian hens and Tasmanian devils are still extant in Tasmania, and the thylacine persisted there until only a century ago. As dingoes colonized Australia but not Tasmania, the dogs have become a major suspect in the extinction of these three creatures from the mainland-they may have preyed upon the hens and outcompeted the other two 33. But some have contested this idea, arguing that this instead had more to do with concurrent human population increase and/or climate change 34.
When it comes to what actually caused those animals to vanish, I don’t have a dog in that fight (pun intended). But it is quite unfortunate that we missed these creatures by a little less than a century and won’t get to see them in the flesh. Regardless, let’s move on from that somber thought to what I think is among the most fascinating aspects of Australian prehistory: the alteration of landscape by the indigenous settlers.
Fire-Stick Farming: Origins and Consequences
The use of fire is one of the most ancient practices in the history of man. Many prehistoric cultures utilized fire not just for cooking and heating, but also to alter the landscapes in which they lived. Australia is perhaps the continent where the link between fire use on the environment by humans is most intimate. Fire-stick farming refers to the controlled use of frequent but low intensity fires by indigenous Australians on landscapes to clear dense vegetation. This helps in promoting fertile grazing areas for prey animals like kangaroos and reducing fuel loads to prevent or mitigate large high intensity wildfires during the dry season.

But exactly when human-driven fire became a prominent character of Australian landscapes, and what exact effects this had, has been a source of contention. Determining the extent and influence of human-induced fire in Australia is not as straightforward as it might seem on the surface. A number of confounding factors make this difficult, such as the loss of megafauna (who may have kept fire in check to some extent), the cold temperatures and low-biomass for much of the Late Pleistocene which would have dampened fire frequency and intensity, changes in human population density, and variation in the sensitivity of different ecosystems to fire.
There are some who argue that humans intensified the fire regime in Australia and that this had huge, often negative consequences for flora and fauna 35. Others have rejected the idea that fire had a major impact, arguing instead that periodically intense fire regimes were a natural part of Australia’s ecological history and variability was controlled mainly by climate 36. Disentangling the influence of natural climate change and humans in shaping fire dynamics and vegetation necessarily requires carefully analyzing long records of pollen, charcoal, and chemicals covering multiple climate cycles and several regions.
The Evidence
Let’s provide some important background on flora: plants in Australasia are often divided into rainforest and sclerophyllous types in the scientific literature. The “rainforest” vegetation is a relict of the ancient and once widespread Gondwanan rainforests. Despite the name, it’s not restricted solely to true rainforests and can be found in moist climates throughout Australasia generally, and it is sensitive to fire. Sclerophyllous vegetation, on the other hand, proliferated over the past several million years in response to the increasingly arid and fire-prone environments in Australia. The wooded areas of Australia are today mostly dominated by sclerophylls (Eucalyptus being most prominent), as they are well-adapted to the mostly dry climate and frequent fires there.


Right: The ancient Daintree Rainforest in, Queensland, northeast Australia. A relict of the Gondwanan rainforests
Increasingly, it seems that human firing of landscapes was prominent, and that this did have noticeable impacts on floral composition in certain areas (the impact on fauna is much less clear, it should be said). The most striking change associated with the onset of burning was a substantial decrease in the proportion of rainforest flora and concomitant increase in the proportion of sclerophylls in areas where the two were competitive with each other-i.e. one was not strongly favored over the other by climate conditions.
Lynch’s Crater in northeast Australia records a major increase in charcoal concentration at around 40 thousand years ago, indicating a surge in fire 37 38. The vegetation just prior to this fire increase was a forest with mixed rainforest and sclerophyll trees. Afterwards, rainforest gymnosperm and angiosperm plants began to decline to a level that was unprecedented during the prior ~200 thousand or so years 37. Rainforest taxa had always had a healthy representation in the Lynch flora record even through the penultimate glacial maximum, but were almost entirely replaced by sclerophyll taxa between about 30 and 12 thousand years ago, only recovering during the Holocene.
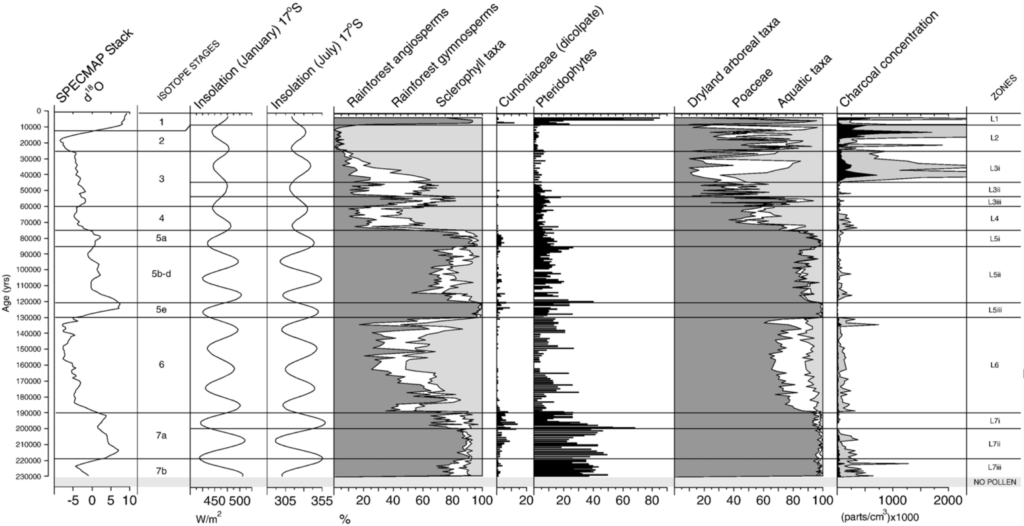
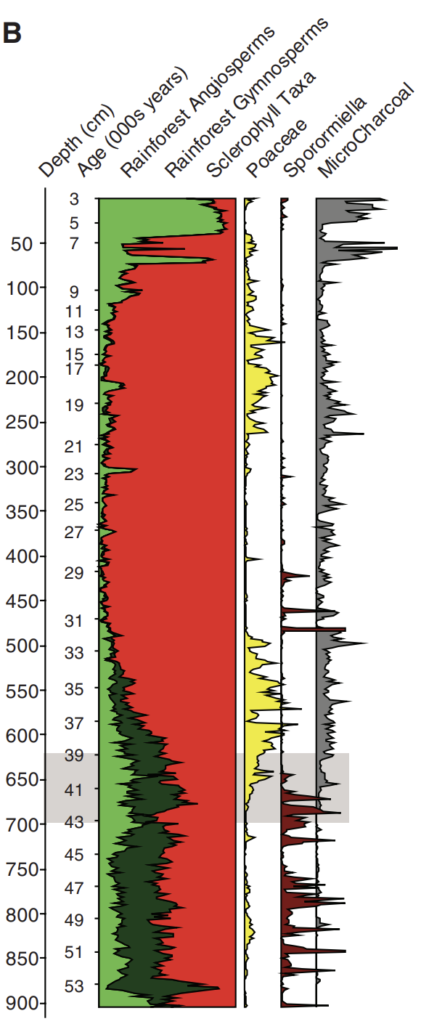
The most parsimonious explanation here is that the increase in fire allowed sclerophyll trees to outcompete rainforest trees, at least for a lengthy period of time. But one study attempts to dispute the notion that the Araucaria (a type of evergreen conifer and one of the “rainforest” taxa) decline at Lynch’s Crater was driven by human-induced fire by revealing a roughly contemporaneous decline in Araucaria pollen at Lake Xere Wapo on the island territory of New Caledonia, where humans did not arrive until only a few thousand years ago and where fire activity was low compared to Lynch’s Crater during the same interval 39.
But the crucial difference is that at Lake Xere Wapo, moist-climate plants of a different variety continued to dominate whereas at Lynch’s Crater, they were almost fully replaced by sclerophyll taxa following human arrival and the surge in fire. Hence, the similarity in the Araucaria decline in both locales says nothing, and in fact the overall pollen and charcoal record actually bolsters the idea that human burning drastically altered the vegetation at Lynch’s Crater by creating a competitive advantage for the sclerophyll trees over the rainforest trees.
At Girraween lagoon near the coast of northern Australia, scientists used geochemical analysis to determine that a human-driven fire regime could have been in place as early as 30 thousand years ago 40. The scientists suspected that anthropogenic fires would have different chemical signatures from natural ones. Human-started fires would be frequent but low intensity and cooler, similar to known fire-stick farming practices in more recent times, whereas natural fires would be rarer but higher intensity (and hotter). The former would result in a larger number of charcoal particles, whereas the latter would burn hotter, resulting in higher values of the geochemical proxy known as MARSPAC.
They analyzed how these chemical signatures changed over time to see if there was a difference in the nature of the fire regime between time periods when humans were present vs. when they weren’t. And remarkably, this is exactly what we see. The number of charcoal particles for the Holocene was around double that for the previous warm, heavily wooded periods during MIS 5 when humans were absent. Meanwhile, the MARSPAC of MIS 2 (which includes the Last Glacial Maximum) was about one third that of MIS 6 (the penultimate Glacial Maximum), a result of the less intense but more frequent burning of lands in the former as a result of human activity.
At both Lynch’s Crater and Girraween lagoon, charcoal levels were not as high during the Last Glacial Maximum as they were during the Holocene. Much of this was obviously due to the greater fuel load during the Holocene when there was more vegetative biomass due to ameliorated climate conditions. But also, Bird et al. 2024 speculate that humans, who may have been developing land-burning techniques for millennia on the now submerged northern continental shelf, moved further inland due to sea level rise after the LGM 40. The same may have applied in the area around Lynch’s Crater, though the change in exposed shelf area was not as large in the northeast as in the north. Hence, an increased population in these regions, combined with higher biomass availability during the Holocene, could have led to the sharp increase in fire.
Moving beyond mainland Australia, the first people to settle Tasmania also seem to have burned through glacial rainforest vegetation in relatively wet areas, in this case causing a floral shift towards heathland 41. The drier heathland regions of the island do not appear to have been as strongly affected, with a few fire-sensitive taxa disappearing but overall no major vegetative turnover. This is a crucial point, as we’ll discuss how geographical/habitat variation in sensitivity to fire plays into all of this later.
Patterns and Implications
Clearly both climate and humans impacted the fire-regime in Australia, with anthropogenic influence being superimposed on broader climate-driven trends. But we might consider that the effects of burning during the Holocene were not the same as during the glacial period. During glacial periods, trees struggle to grow due to low CO2 and as a result, saplings may not reach a fire-proof size in time to withstand fires 42. This would normally be offset by the fact that the rarity of lightning strikes due to fewer thunderstorms, low temperatures, and low biomass during the coldest of glacial times reduced the likelihood of fire ignition and decreased fire intensity.
However, with humans on the scene as an additional source of ignition, and more specifically frequent ignitions, already stressed glacial trees (or at least certain types that were not fire-adapted) could have been dealt a severe blow. This may explain why at Lynch’s Crater, rainforest tree taxa were drastically reduced in abundance between roughly 30 and 12 thousand years ago when harsh glacial conditions and human-induced fire were both present. It could also explain how they were prominent during the Holocene when the fire regime was strong but conditions were more conducive for tree growth 38, and also had a healthy representation during the penultimate glacial maximum which was low-productivity but also low fire 37.
The climate of Australia during the Last Glacial Maximum around 21 thousand years ago continues to be debated. Some more recent papers have bucked traditional notions of ice age aridity by arguing that sections of glacial periods, including the LGM, could’ve been wetter than once thought 43 44. They’ve suggested that the apparent discrepancy with this purportedly wet climate and LGM pollen data showing more open, drier vegetation compared to the Holocene may be reconcilable with the effects of low CO2 on vegetation, which is thought to increase water stress by reducing plant water use efficiency.
The possibility that human fire-use shaped LGM vegetation was not considered in those studies. It could well be that a complex synergistic interaction between climate (namely lower temperatures), low CO2, and human fire-use could better explain LGM vegetation in Australia than low CO2 alone. The disproportionate impact of anthropogenic fire on Australian vegetation in certain regions like Lynch’s Crater or parts of Tasmania may be partially explained by its use during ecologically sensitive intervals.
Still, not all regions experienced a major vegetative shift, such as the Murray-Darling basin in southeast Australia which had a dry climate and essentially savannah-dominated environment both prior to and following the onset of anthropogenic burning 45. The dry heathland-dominated areas of Tasmania also did not experience a strong floral turnover unlike the forested ones 41. In New Guinea, one of the wettest places in the world, there is evidence of increased charcoal accumulation- potentially associated with human activity-during the Late Pleistocene 46. However, there’s no evidence for a substantial decline in rainforest cover, at least until the Holocene when land-clearance associated with agriculture became prominent there 47.

The available information so far seems to point towards there being some sort of “sweet spot” with regard to how strongly Sahul’s ecosystems were impacted by human-caused fire, with areas that were neither too dry nor excessively wet being the most affected (i.e. moderately wet places). Dry areas like the savannah of Murray Darling already contained fire-resistant vegetation and/or had low fuel availability, whereas New Guinea could’ve been too moist for Pleistocene foragers to effectively start and maintain fires with the meager tools at their disposal. Population density is another confounding factor-New Guinea clearly had a very small population for a while as evidenced by genetic bottlenecks and slow population growth 7.
There’s also been some tantalizing speculation that the introduction of an anthropogenic fire regime into Australia may have even altered the climate. The story goes that the reduction of vegetation, particularly trees, through burning reduced evapotranspiration and hence rainfall in downwind, interior regions 48 49. This allegedly caused inland lakes like Kati Thanda (Lake Eyre) to remain dry from around 50 thousand years ago until the present, even at times when the monsoon should have theoretically been strong-unique in the Quaternary. This idea is currently controversial with other papers either contesting the conclusions or arguing that the evidence is inconclusive 50 51.
More research is needed, but if it’s true, it could represent one of the earliest examples of humans impacting climate. Regardless, there’s enough evidence that humans had profound impacts on their landscapes, and the fact that this occurred in deep time and involved people living at low densities with only stone age technology is a huge part of why this topic is so intriguing to me.
Fire: All Bad?
At this point, it may appear that fire-stick farming as a practice is conclusively harmful, as ecosystems were often impacted in decisive, potentially negative ways. But paradoxically, many have argued for the benefits of this practice. Recall how we started this section by mentioning how frequent controlled fires are suggested to improve foraging grounds for animals and, more importantly, reduce the likelihood of damaging high intensity fires occurring. The increase in extreme fire events in post-colonization after which the indigenous land management technique was largely discontinued, is considered evidence for this 52.
During the Pleistocene, Australian megafauna are speculated to have at least partly blunted fire intensity by consuming copious amounts of vegetation and hence reducing the fuel load 51 53. Those animals are no longer with us, meaning that strategic use of low-intensity fire as practiced by some aboriginal groups may be a good option to mitigate the sorts of severe wildfire events that characterize Australia during warm interglacial periods, like our current one.
Last Remarks
Australia is a place that continues to fascinate me, even though I’m yet to go there. This post about Sahul, the ancient landmass that included not just mainland Australia but also Tasmania and New Guinea, is one I’ve been wanting to write for a while. I was inspired to do so after seeing how fraught with misconception subjects like the settlement of Sahul and the origins of its native people are, and also to tell the story of other things that I’ve researched, like fire-stick farming.
There’s a lot that still needs to be resolved, of course. For example, what impact dingoes actually had on native species, when Denisovan introgression into Oceanians actually occurred, and what the true effects of a human fire regime on Australia were. With more studies, we’ll get a clearer picture. At the moment though, I think we should steer clear of ideas that aren’t backed up by evidence, however enticing they may appear.
References
- Clarkson, C. (2017). Human occupation of northern Australia by 65,000 years ago. Nature, 547(7663), 306–310. https://doi.org/10.1038/nature22968[↩]
- Westaway, K. E., Louys, J., Awe, R. D., Morwood, M. J., Price, G. J., Zhao, J., Aubert, M., Joannes-Boyau, R., Smith, T. M., Skinner, M. M., Compton, T., Bailey, R. M., van den Bergh, G. D., de Vos, J., Pike, A. W. G., Stringer, C., Saptomo, E. W., Rizal, Y., Zaim, J., & Santoso, W. D. (2017). An early modern human presence in Sumatra 73,000–63,000 years ago. Nature, 548(7667), 322–325. https://doi.org/10.1038/nature23452[↩]
- Freidline, S. E., Westaway, K. E., Joannes-Boyau, R., Duringer, P., Ponche, J.-L., Morley, M. W., Hernandez, V. C., McAllister-Hayward, M. S., McColl, H., Zanolli, C., Gunz, P., Bergmann, I., Sichanthongtip, P., Sihanam, D., Boualaphane, S., Luangkhoth, T., Souksavatdy, V., Dosseto, A., Boesch, Q., & Patole-Edoumba, E. (2023). Early presence of Homo sapiens in Southeast Asia by 86–68 kyr at Tam Pà Ling, Northern Laos. Nature Communications, 14(1), 3193. https://doi.org/10.1038/s41467-023-38715-y[↩]
- O’Connell, J. F., Allen, J., Williams, M. A. J., Williams, A. N., Turney, C. S. M., Spooner, N. A., Kamminga, J., Brown, G., & Cooper, A. (2018). When did Homo sapiens first reach Southeast Asia and Sahul? Proceedings of the National Academy of Sciences, 115(34), 8482–8490. https://doi.org/10.1073/pnas.1808385115[↩]
- Williams, M. A. J., Spooner, N. A., McDonnell, K., & O’Connell, J. F. (2020). Identifying disturbance in archaeological sites in tropical northern Australia: Implications for previously proposed 65,000‐year continental occupation date. Geoarchaeology, 36(1), 92–108. https://doi.org/10.1002/gea.21822[↩]
- Tobler, R., Rohrlach, A., Soubrier, J., Bover, P., Llamas, B., Tuke, J., Bean, N., Abdullah-Highfold, A., Agius, S., O’Donoghue, A., O’Loughlin, I., Sutton, P., Zilio, F., Walshe, K., Williams, A. N., Turney, C. S. M., Williams, M., Richards, S. M., Mitchell, R. J., & Kowal, E. (2017). Aboriginal mitogenomes reveal 50,000 years of regionalism in Australia. Nature, 544(7649), 180–184. https://doi.org/10.1038/nature21416[↩][↩][↩]
- Mondal, M., André, M., Pathak, A. K., Brucato, N., Ricaut, F.-X., Metspalu, M., & Eriksson, A. (2025). Resolving out of Africa event for Papua New Guinean population using neural network. Nature Communications, 16(1). https://doi.org/10.1038/s41467-025-61661-w[↩][↩][↩][↩][↩][↩]
- Allen, J., & O’Connell, J. F. (2025). Recent DNA Studies Question a 65 kya Arrival of Humans in Sahul. Archaeology in Oceania/Archæology & Physical Anthropology in Oceania, 60(2), 187–190. https://doi.org/10.1002/arco.70002[↩]
- Rasmussen, M., Guo, X., Wang, Y., Lohmueller, K. E., Rasmussen, S., Albrechtsen, A., Skotte, L., Lindgreen, S., Metspalu, M., Jombart, T., Kivisild, T., Zhai, W., Eriksson, A., Manica, A., Orlando, L., Vega, F. M. D. L., Tridico, S., Metspalu, E., Nielsen, K., & Ávila-Arcos, M. C. (2011). An Aboriginal Australian Genome Reveals Separate Human Dispersals into Asia. Science, 334(6052), 94–98. https://doi.org/10.1126/science.1211177[↩]
- Mallick, S., Li, H., Lipson, M., Mathieson, I., Gymrek, M., Racimo, F., Zhao, M., Chennagiri, N., Nordenfelt, S., Tandon, A., Skoglund, P., Lazaridis, I., Sankararaman, S., Fu, Q., Rohland, N., Renaud, G., Erlich, Y., Willems, T., Gallo, C., & Spence, J. P. (2016). The Simons Genome Diversity Project: 300 genomes from 142 diverse populations. Nature, 538(7624), 201–206. https://doi.org/10.1038/nature18964[↩]
- Lipson, M., & Reich, D. (2017). A working model of the deep relationships of diverse modern human genetic lineages outside of Africa. Molecular Biology and Evolution, msw293. https://doi.org/10.1093/molbev/msw293[↩][↩]
- Pagani, L., Lawson, D. J., Jagoda, E., Mörseburg, A., Eriksson, A., Mitt, M., Clemente, F., Hudjashov, G., DeGiorgio, M., Saag, L., Wall, J. D., Cardona, A., Mägi, R., Wilson Sayres, M. A., Kaewert, S., Inchley, C., Scheib, C. L., Järve, M., Karmin, M., & Jacobs, G. S. (2016). Genomic analyses inform on migration events during the peopling of Eurasia. Nature, 538(7624), 238–242. https://doi.org/10.1038/nature19792[↩]
- Sümer, A. P., Rougier, H., Villalba-Mouco, V., Huang, Y., Leonardo, Essel, E., Mesa, A. B., Furtwaengler, A., Stéphane Peyrégne, Filippo, C. de, Rohrlach, A. B., Pierini, F., Fabrizio Mafessoni, Fewlass, H., Zavala, E. I., Mylopotamitaki, D., Bianco, R. A., Schmidt, A., Zorn, J., & Nickel, B. (2024). Earliest modern human genomes constrain timing of Neanderthal admixture. Nature. https://doi.org/10.1038/s41586-024-08420-x[↩]
- Prüfer, K., Racimo, F., Patterson, N., Jay, F., Sankararaman, S., Sawyer, S., Heinze, A., Renaud, G., Sudmant, P. H., de Filippo, C., Li, H., Mallick, S., Dannemann, M., Fu, Q., Kircher, M., Kuhlwilm, M., Lachmann, M., Meyer, M., Ongyerth, M., & Siebauer, M. (2013). The complete genome sequence of a Neanderthal from the Altai Mountains. Nature, 505(7481), 43–49. https://doi.org/10.1038/nature12886[↩][↩]
- Slon, V., Mafessoni, F., Vernot, B., de Filippo, C., Grote, S., Viola, B., Hajdinjak, M., Peyrégne, S., Nagel, S., Brown, S., Douka, K., Higham, T., Kozlikin, M. B., Shunkov, M. V., Derevianko, A. P., Kelso, J., Meyer, M., Prüfer, K., & Pääbo, S. (2018). The genome of the offspring of a Neanderthal mother and a Denisovan father. Nature, 561(7721), 113–116. https://doi.org/10.1038/s41586-018-0455-x[↩]
- Reich, D., Patterson, N., Kircher, M., Delfin, F., Nandineni, Madhusudan R., Pugach, I., Ko, A.-S., Ko, Y.-C., Jinam, Timothy A., Phipps, Maude E., Saitou, N., Wollstein, A., Kayser, M., Pääbo, S., & Stoneking, M. (2011). Denisova Admixture and the First Modern Human Dispersals into Southeast Asia and Oceania. The American Journal of Human Genetics, 89(4), 516–528. https://doi.org/10.1016/j.ajhg.2011.09.005[↩][↩]
- Stringer, C. B., & Barnes, I. (2015). Deciphering the Denisovans. Proceedings of the National Academy of Sciences, 112(51), 15542–15543. https://doi.org/10.1073/pnas.1522477112[↩]
- Sankararaman, S., Mallick, S., Patterson, N., & Reich, D. (2016). The Combined Landscape of Denisovan and Neanderthal Ancestry in Present-Day Humans. Current Biology, 26(9), 1241–1247. https://doi.org/10.1016/j.cub.2016.03.037[↩]
- Jacobs, G. S., Hudjashov, G., Saag, L., Kusuma, P., Darusallam, C. C., Lawson, D. J., Mondal, M., Pagani, L., Ricaut, F.-X., Stoneking, M., Metspalu, M., Sudoyo, H., Lansing, J. S., & Cox, M. P. (2019). Multiple Deeply Divergent Denisovan Ancestries in Papuans. Cell, 177(4), 1010-1021.e32. https://doi.org/10.1016/j.cell.2019.02.035[↩]
- Pedro, N., Brucato, N., Fernandes, V., André, M., Saag, L., Pomat, W., Besse, C., Boland, A., Deleuze, J.-F., Clarkson, C., Sudoyo, H., Metspalu, M., Stoneking, M., Cox, M. P., Leavesley, M., Pereira, L., & Ricaut, F.-X. (2020). Papuan mitochondrial genomes and the settlement of Sahul. Journal of Human Genetics, 65(10), 875–887. https://doi.org/10.1038/s10038-020-0781-3[↩]
- Huxley, T. H. (1870). On the Geographical Distribution of the Chief Modifications of Mankind. The Journal of the Ethnological Society of London (1869-1870), 2(4), 404. https://doi.org/10.2307/3014371[↩]
- Redd, A. J., Roberts-Thomson, J., Karafet, T., Bamshad, M., Jorde, L. B., Naidu, J. M., Walsh, B., & Hammer, M. F. (2002). Gene Flow from the Indian Subcontinent to Australia: Evidence from the Y Chromosome. Current Biology, 12(8), 673–677. https://doi.org/10.1016/S0960-9822(02)00789-3[↩]
- Pugach, I., Delfin, F., Gunnarsdóttir, E., Kayser, M., & Stoneking, M. (2013). Genome-wide data substantiate Holocene gene flow from India to Australia. Proceedings of the National Academy of Sciences, 110(5), 1803–1808. https://doi.org/10.1073/pnas.1211927110[↩]
- Hudjashov, G., Kivisild, T., Underhill, P. A., Endicott, P., Sanchez, J. J., Lin, A. A., Shen, P., Oefner, P., Renfrew, C., Villems, R., & Forster, P. (2007). Revealing the prehistoric settlement of Australia by Y chromosome and mtDNA analysis. Proceedings of the National Academy of Sciences, 104(21), 8726–8730. https://doi.org/10.1073/pnas.0702928104[↩]
- Bergström, A., Nagle, N., Chen, Y., McCarthy, S., Pollard, M. O., Ayub, Q., … & Tyler-Smith, C. (2016). Deep roots for Aboriginal Australian Y chromosomes. Current Biology, 26(6), 809-813.[↩]
- Nagle, N., Ballantyne, K. N., van Oven, M., Tyler-Smith, C., Xue, Y., Taylor, D., Wilcox, S., Wilcox, L., Turkalov, R., van Oorschot, R. A. H., McAllister, P., Williams, L., Kayser, M., & Mitchell, R. J. (2015). Antiquity and diversity of aboriginal Australian Y-chromosomes. American Journal of Physical Anthropology, 159(3), 367–381. https://doi.org/10.1002/ajpa.22886[↩]
- Nagle, N., Ballantyne, K. N., van Oven, M., Tyler-Smith, C., Xue, Y., Wilcox, S., Wilcox, L., Turkalov, R., van Oorschot, R. A. H., van Holst Pellekaan, S., Schurr, T. G., McAllister, P., Williams, L., Kayser, M., & Mitchell, R. J. (2016). Mitochondrial DNA diversity of present-day Aboriginal Australians and implications for human evolution in Oceania. Journal of Human Genetics, 62(3), 343–353. https://doi.org/10.1038/jhg.2016.147[↩]
- Vallini, L., Zampieri, C., Shoaee, M. J., Bortolini, E., Marciani, G., Aneli, S., Pievani, T., Benazzi, S., Barausse, A., Mezzavilla, M., Petraglia, M. D., & Pagani, L. (2024). The Persian plateau served as hub for Homo sapiens after the main out of Africa dispersal. Nature Communications, 15(1), 1882. https://doi.org/10.1038/s41467-024-46161-7[↩]
- Oxenham, M. F., & Buckley, H. (2016). The Routledge handbook of bioarchaeology in Southeast Asia and the Pacific Islands (pp. 471–475). Routledge, Taylor & Francis Group.[↩]
- Fillios, M. A., & Taçon, P. S. C. (2016). Who let the dogs in? A review of the recent genetic evidence for the introduction of the dingo to Australia and implications for the movement of people. Journal of Archaeological Science: Reports, 7, 782–792. https://doi.org/10.1016/j.jasrep.2016.03.001[↩][↩]
- Yassine Souilmi, Wasef, S., Williams, M. P., Conroy, G., Bar, I., Bover, P., Dann, J., Heiniger, H., Llamas, B., Ogbourne, S., Archer, M., Ballard, W. O., Reed, E., Tobler, R., Loukas Koungoulos, Walshe, K., Wright, J. L., Balme, J., O’Connor, S., & Cooper, A. (2024). Ancient genomes reveal over two thousand years of dingo population structure. Proceedings of the National Academy of Sciences, 121(30). https://doi.org/10.1073/pnas.2407584121[↩][↩]
- Gollan, K. (1985). Prehistoric Dogs in Australia: An Indian Origin? Recent Advances in Indo-Pacific Prehistory, 439–443. https://doi.org/10.1163/9789004644472_064[↩]
- Johnson, C. N., & Wroe, S. (2003). Causes of extinction of vertebrates during the Holocene of mainland Australia: arrival of the dingo, or human impact? The Holocene, 13(6), 941–948. https://doi.org/10.1191/0959683603hl682fa[↩]
- Prowse, T. A. A., Johnson, C. N., Bradshaw, C. J. A., & Brook, B. W. (2014). An ecological regime shift resulting from disrupted predator–prey interactions in Holocene Australia. Ecology, 95(3), 693–702. https://doi.org/10.1890/13-0746.1[↩]
- Miller, G. H. (2005). Ecosystem Collapse in Pleistocene Australia and a Human Role in Megafaunal Extinction. Science, 309(5732), 287–290. https://doi.org/10.1126/science.1111288[↩]
- Mooney, S. D., Harrison, S. P., Bartlein, P. J., Daniau, A.-L. ., Stevenson, J., Brownlie, K. C., Buckman, S., Cupper, M., Luly, J., Black, M., Colhoun, E., D’Costa, D., Dodson, J., Haberle, S., Hope, G. S., Kershaw, P., Kenyon, C., McKenzie, M., & Williams, N. (2011). Late Quaternary fire regimes of Australasia. Quaternary Science Reviews, 30(1), 28–46. https://doi.org/10.1016/j.quascirev.2010.10.010[↩]
- Kershaw, A. P., Bretherton, S. C., & van der Kaars, S. (2007). A complete pollen record of the last 230 ka from Lynch’s Crater, north-eastern Australia. Palaeogeography, Palaeoclimatology, Palaeoecology, 251(1), 23–45. https://doi.org/10.1016/j.palaeo.2007.02.015[↩][↩][↩]
- Rule, S., Brook, B. W., Haberle, S. G., Turney, C. S. M., Kershaw, A. P., & Johnson, C. N. (2012). The Aftermath of Megafaunal Extinction: Ecosystem Transformation in Pleistocene Australia. Science, 335(6075), 1483–1486. https://doi.org/10.1126/science.1214261[↩][↩]
- Stevenson, J., & Hope, G. (2005). A Comparison of late Quaternary Forest Changes in New Caledonia and Northeastern Australia. Quaternary Research, 64(3), 372–383. https://doi.org/10.1016/j.yqres.2005.08.011[↩]
- Bird, M. I., Brand, M., Comley, R., Fu, X., Hadeen, X., Jacobs, Z., Rowe, C., Wurster, C. M., Zwart, C., & Bradshaw, C. J. A. (2024). Late Pleistocene emergence of an anthropogenic fire regime in Australia’s tropical savannahs. Nature Geoscience, 17(3), 233–240. https://doi.org/10.1038/s41561-024-01388-3[↩][↩]
- Adeleye, M. A., Hopf, F., Haberle, S. G., Stannard, G. L., Mcwethy, D. B., Harris, S., & David. (2024). Landscape burning facilitated Aboriginal migration into Lutruwita/Tasmania 41,600 years ago. Science Advances, 10(46). https://doi.org/10.1126/sciadv.adp6579[↩][↩]
- Bond, W. J., Midgley, G. F., & Woodward, F. I. (2003). The importance of low atmospheric CO2 and fire in promoting the spread of grasslands and savannas. Global Change Biology, 9(7), 973–982. https://doi.org/10.1046/j.1365-2486.2003.00577.x[↩]
- Weij, R., Sniderman, J. M. K., Woodhead, J. D., Hellstrom, J. C., Brown, J. R., Drysdale, R. N., Reed, E., Bourne, S., & Gordon, J. (2024). Elevated Southern Hemisphere moisture availability during glacial periods. Nature, 626(7998), 319–326. https://doi.org/10.1038/s41586-023-06989-3[↩]
- Cadd, H., Williams, A. N., Saktura, W. M., Cohen, T. J., Mooney, S. D., He, C., Otto‐Bliesner, B., & Chris. (2024). Last Glacial Maximum cooling induced positive moisture balance and maintained stable human populations in Australia. Communications Earth & Environment, 5(1). https://doi.org/10.1038/s43247-024-01204-1[↩]
- Adeleye, M. A., Charles Andrew, S., Gallagher, R., van der Kaars, S., De Deckker, P., Hua, Q., & Haberle, S. G. (2023). On the timing of megafaunal extinction and associated floristic consequences in Australia through the lens of functional palaeoecology. Quaternary Science Reviews, 316, 108263. https://doi.org/10.1016/j.quascirev.2023.108263[↩]
- Summerhayes, G. R., Field, J. H., Shaw, B., & Gaffney, D. (2017). The archaeology of forest exploitation and change in the tropics during the Pleistocene: The case of Northern Sahul (Pleistocene New Guinea). Quaternary International, 448, 14–30. https://doi.org/10.1016/j.quaint.2016.04.023[↩]
- Haberle, S. G., Lentfer, C., O’Donnell, S., & Denham, T. (2012). The palaeoenvironments of Kuk Swamp from the beginnings of agriculture in the highlands of Papua New Guinea. Quaternary International, 249, 129–139. https://doi.org/10.1016/j.quaint.2011.07.048[↩]
- Miller, G., Mangan, J., Pollard, D., Thompson, S., Felzer, B., & Magee, J. (2005). Sensitivity of the Australian Monsoon to insolation and vegetation: Implications for human impact on continental moisture balance. Geology, 33(1), 65. https://doi.org/10.1130/g21033.1[↩]
- Miller, G. H., Fogel, M. L., Magee, J. W., & Gagan, M. K. (2016). Disentangling the impacts of climate and human colonization on the flora and fauna of the Australian arid zone over the past 100 ka using stable isotopes in avian eggshell. Quaternary Science Reviews, 151, 27–57. https://doi.org/10.1016/j.quascirev.2016.08.009[↩]
- Cohen, T. J., Arnold, L. J., Gázquez, F., May, J.-H., Marx, S. K., Jankowski, N. R., Chivas, A. R., Garćia, A., Cadd, H., Parker, A. G., Jansen, J. D., Fu, X., Waldmann, N., Nanson, G. C., Jones, B. G., & Gadd, P. (2022). Late Quaternary climate change in Australia’s arid interior: Evidence from Kati Thanda – Lake Eyre. Quaternary Science Reviews, 292, 107635. https://doi.org/10.1016/j.quascirev.2022.107635[↩]
- Bird, M. I., Hutley, L. B., Lawes, M. J., Lloyd, J., Luly, J. G., Ridd, P. V., Roberts, R. G., Ulm, S., & Wurster, C. M. (2013). Humans, megafauna and environmental change in tropical Australia. Journal of Quaternary Science, 28(5), 439–452. https://doi.org/10.1002/jqs.2639[↩][↩]
- Mariani, M., Connor, S. E., Theuerkauf, M., Herbert, A., Kuneš, P., Bowman, D., Fletcher, M., Head, L., Kershaw, A. P., Haberle, S. G., Stevenson, J., Adeleye, M., Cadd, H., Hopf, F., & Briles, C. (2022). Disruption of cultural burning promotes shrub encroachment and unprecedented wildfires. Frontiers in Ecology and the Environment, 20(5), 292–300. https://doi.org/10.1002/fee.2395[↩]
- Flannery, T. F. (1990). Pleistocene faunal loss: implications of the aftershock for Australia’s past and future. Archaeology in Oceania, 25(2), 45–55. https://doi.org/10.1002/j.1834-4453.1990.tb00232.x[↩]