The mammoth steppe. Anyone who has heard the term is guaranteed to be at least somewhat familiar with Pleistocene paleontology and paleoecology. It was the largest biome in the world during ice ages, stretching from the Atlantic Ocean in Western Europe all the way through Siberia and into Alaska and the Yukon, which has made it the subject of intense research by scientists and fascination by prehistory enthusiasts.
I’ve already written about the mammoth steppe extensively on this very blog, with a large number of posts referencing it. But there is still a great deal to discuss, and this article will hopefully fill in some of the gaps left by my earlier posts. Today’s rather lengthy article will first give a brief background on what the mammoth steppe was and how it formed, and then go into a much longer discussion about how it, as well as much of its resident fauna, disappeared.
I have not gone into depth on the Late Pleistocene extinctions since last year, but it is necessary to do so when discussing this particular topic. Hence, this article will include a deep and thorough analysis of the paleoecological and archeological context that surrounds the demise of these animals, principally on the Eurasian mainland.
We will also address to what extent the mammoth steppe can be “restored” and what current Pleistocene rewilding efforts in Arctic and Subarctic regions can realistically hope to accomplish.
With that said, let’s move to the overview.
Overview of the Mammoth Steppe (and Controversies)
The term mammoth steppe was originally coined by the late R. Dale Guthrie, one of the world’s foremost experts on this paleo-biome1. Mammoth steppe is often, but not always, used synonymously with steppe-tundra or tundra-steppe. For the purpose of this article, I will use them interchangeably. It was an environment characterized by cold and typically dry climates, although the extent of cold and aridity obviously differed greatly depending on the region. Trees were usually scarce or absent.
Mammoth steppe or steppe-tundra is distinct from modern tundras in that plants that are associated with steppes such as graminoids (grasses and sedges) and forbs (especially Artemisia) were far more prominent in the former than the latter, which has relatively higher concentration of shrubs and mosses. It differed from temperate steppes in that plants associated with Arctic climates such as dwarf shrubs of the birch and willow families and the Dryas flower (avens) were also present. Rather than being a homogeneous block, it was more likely a mosaic of habitats with some patches being more steppe-like and others more tundra-like2. Its distribution fluctuated through time and it was bounded by oceans, woodlands, cold deserts, and ice sheets.
The inherent allure of the mammoth steppe is partly due to the difficulty in imagining vast grassy plains existing in place of what are now dense boreal forests. But the biggest source of intrigue with this ecosystem is probably the creatures it hosted: the famous woolly mammoth, woolly rhino, steppe bison, horse, reindeer, saiga antelope, cave lion and, in some areas, cave hyena as well as numerous smaller fauna. These animals are collectively referred to as the Mammuthus-Coelodonta faunal complex3, named after the woolly mammoth and woolly rhinoceros who were key components.

The sheer number of large animal remains unearthed from areas of the former mammoth steppe indicates that it was still productive despite the cold climate. The key to said productivity is understood to be the abundance of nutritious herbaceous vegetation such as grasses and forbs 4 5 6, which is more ideal for grazers than the current vegetation that is nutrient-poor and contains anti-herbivory chemical compounds.
Only a few isolated analogs of this ecosystem exist today. According to many scientists, such as Dale Guthrie, climate change at the end of the Pleistocene was responsible for its loss as wetter conditions coincident with warming made it difficult to maintain the “steppic” profile of the mammoth steppe, causing it to eventually become tundra north of the Arctic treeline, and boreal forest south of it 4. Guthrie endorses a common position that the vanishing of the mammoth steppe then made it impossible to support the large herds of grazers that once lived there, as the herbaceous vegetation was replaced by less palatable shrubs and trees.
However, whether the disappearance of the mammoth steppe caused an extinction of its megafauna or if the extinction of the megafauna caused a disappearance of the mammoth steppe is a matter of heated debate. Russian scientist Sergey Zimov has been a champion of the latter concept, arguing it was actually human-driven demise of Pleistocene megafauna through overhunting that caused this transformation via top-down mechanisms 7 8. Without herbivores, it is said, nutrients were not cycled properly as there were no large herds of animals eating plants and releasing waste, and shrubs and trees could grow unimpeded as they were no longer being trampled.

Zimov has gone as far as to create a park in northern Siberia stocked with introduced herbivores in an attempt to show that the mammoth steppe’s disappearance was not natural and that it can be restored with Pleistocene rewilding 9. He also believes that the extinction of the fauna and the top-down effects it had on vegetation reduced the Earth’s capacity to maintain permafrost and prevent carbon from seeping into the atmosphere, a dimension that has garnered attention as the Earth warms in modern times and there is concern of permafrost melting and releasing its methane stores into the atmosphere, thereby further amplifying greenhouse warming.
As someone who has heavily researched the topic of the disappearance of the mammoth steppe and the Mammuthus-Coelodonta complex, it needs to be said that the dynamics involved can get confusing and overwhelming. It is far more difficult to disentangle the effects of climate change and humans on the extinction of mammoth steppe fauna than in the case of the New World or Australasian megafauna. Related concepts will need to be decoupled from each other.
At this moment, the large majority of research seems to point to climatically driven losses of both the mammoth steppe and the characteristic fauna 10. Unfortunately, many viewpoints lack nuance and careful consideration of the opposing arguments, with many researchers, particularly those who favor environmental explanations, prematurely jumping to certain conclusions. And although the fates of the mammoth steppe and its faunal complex are intimately and strongly intertwined, I will show that there is reason to partially decouple the two in order to better understand what happened.
The questions we will try to answer here include:
- What parameters allowed the mammoth steppe to exist?
- What caused the disappearance of the mammoth steppe?
- Why did many of the megafauna of the mammoth steppe faunal complex disappear?
- What effects would the megafauna have had on northern ecosystems and what can realistically be achieved with rewilding?
Having established that overview, let’s move on with the topic.
Formation and Contraction
How did the mammoth steppe form? First of all, the mammoth steppe or steppe-tundra sensu stricto was a mostly treeless environment 2 4 . In most cases, the only way that an environment so devoid of trees can occur naturally is if it is too cold (summer temperatures too low to enable tree growth) or too dry (growth of trees inhibited by lack of water), or both. These cold and usually dry conditions were typical of large parts of the northern Holarctic during ice ages.
There were multiple factors that were conducive to the existence of steppic habitats in Arctic and Subarctic climate zones during glacials, where they are largely absent today. Firstly, as Dale Guthrie has pointed out, there were multiple “moisture-blocking” features that reduced precipitation and cloud cover throughout northern latitudes 4. These included a frozen North Atlantic Ocean in winter which reduced evaporation, vastly exposed continental shelves due to lower sea level which increased distance to the sea, and ice sheets that blocked the flow of moisture.
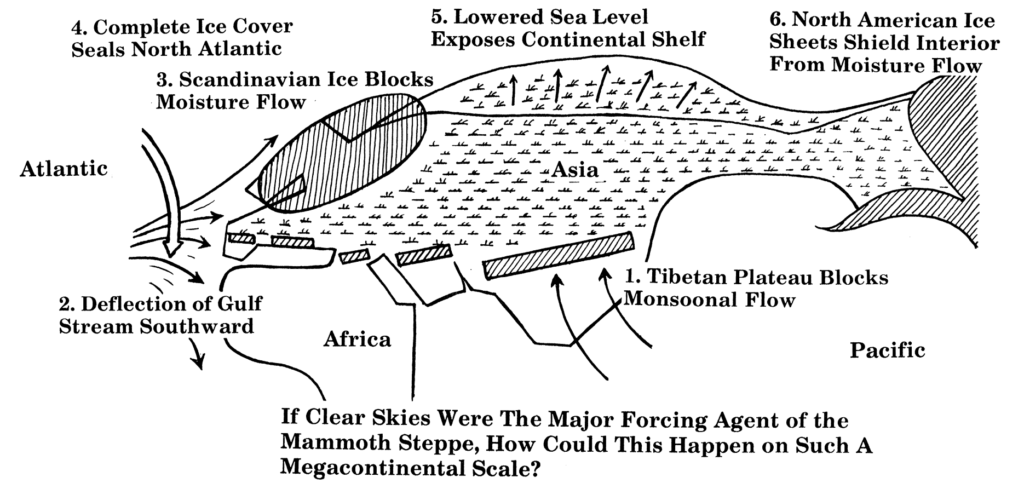
Increased aridity via reduced cloud cover and precipitation had the effect of favoring grasses and forbs over trees and shrubs, as the former require less water. Additionally, the low CO2 of glacial periods may have enhanced this effect by reducing the water-use efficiency of plants11, once again favoring non-woody plants which require less water. Extensive permafrost and colder temperatures shifted the treeline further south in certain areas.
Furthermore, large amounts of loess (wind-blown dust) were deposited over much of the mammoth steppe, which provided essential minerals that fertilized the soil 12. More fertile soils tend to favor herbaceous plants over shrubs and moss. Reduced snowfall had another effect. Snow cover actually allows shrubs to survive by insulating them from very cold winter air temperatures. A thinner snow cover during glacials was therefore bad for shrubs and gave herbaceous plants an advantage as the latter hold most of their biomass underground and can therefore tolerate very cold weather.
Some combination of cold, aridity, and other factors resulted in the disappearance of the boreal forest belt in northern Eurasia, which then allowed plants of the steppe and tundra belts to mix unimpeded3. Hence, an ecosystem distinct from modern ones was formed and stretched from the Atlantic to the Pacific and the Arctic to Central Asia. But even then, there are complexities and uncertainties with regard to the underlying causes of the vegetation structure. For instance, some have suggested that moisture and temperature during the LGM in Europe were too high to inhibit forest growth, and that other factors such as human-induced fire contributed to this openness 13.
Glacial conditions were ideal for the existence of a massive, largely treeless biome covering a vast area of the northern Holarctic. Graminoids and forbs were dominant components of the plant assemblages, and these served as highly nutritious fodder for the large herds of megafauna that roamed the plains. But what happened as the last ice age ended?
This is where things get messy. Increased precipitation due to decaying ice sheets and warmer sea surface temperatures over the North Atlantic resulted in greater overall moisture, which along with the increase in CO2, benefitted shrubs and trees. Smaller dust source areas existed during the Holocene, resulting in less dust/loess reaching the high latitudes 14. Moreover, the warming summers allowed the treeline to shift further north than in typical stadial conditions (trees still grew quite far north in Siberia during interstadials of the last glacial, although they were sparse).
However, the crucial point is that this is not unique to the Holocene, but also occurred in previous interglacials. Pollen data indicates that boreal forests and moist tundra environments of some form or another were widespread during previous interglacials. The surrounding environs of Lake Baikal in southern Siberia featured taiga forest in the Eemian interglacial which had been immediately preceded by a moist shrubby tundra during the late glacial transition period 15, meanwhile the earlier and very lengthy MIS 11 interglacial was also dominated by taiga between 424-396 kya 16. Similarly at Lake Elgygytgyn in Chukotka in northeastern Siberia, the abundance of tundra and/or taiga taxa increased greatly at the expense of steppe taxa going into each interglacial 17.

In eastern Beringia (Alaska and the Yukon), the story is mirrored. In central Alaska during the previous interglacial, boreal forest was extensive 18. In the Yukon, boreal forests during previous interglacials had a different composition in comparison to the late Holocene in terms of tree types with more spruce and fir and less pine 19, but were still forest at the end of the day. This is despite the fact that permafrost-which can act as a source of moisture-was reduced as a result of warmer summers. Increased precipitation probably compensated, resulting in conditions as wet or wetter than the present.
The big takeaway here is that the pollen data is problematic for both camps in the “mammoth steppe” debate. For those that argue the mammoth steppe was maintained by the herbivores, the evidence suggests tree/shrub cover comparable to that of the Holocene dominating during interglacials in key regions, possibly indicating the effects of the animals on vegetation may have been minimal and/or localized. On the other hand, those who argue that climate was the cause of the disappearance of the mammoth steppe and its megafauna would struggle to explain why the mammoth steppe fauna persevered during past interglacials but not the current one despite the former not differing much vegetatively from the latter.
This may be somewhat confusing and leads us to the topic of how mammoth steppe environmental changes relate to extinction.
The Holocene Exception?
One of the key questions with regard to the extinction of steppe-tundra megafauna, just like with the extinction of megafauna during the Late Pleistocene and early Holocene in general, is why these animals survived previous climate cycles but not the most recent one. We do not know the exact distribution of woolly mammoths, woolly rhinos, steppe bison, and others during previous interglacials but we obviously do know that they survived them. The question is how?
For those who argue for climatic causes, this dilemma is difficult to solve but explanations have been attempted. Namely, supporters of this viewpoint tend to argue that something truly was/is unique about the Holocene from a climatic perspective. It was once claimed that the Eemian was less stable than the current interglacial with “flickering” of the climate between warm and cold, resulting in a perpetually frozen Arctic Ocean, which in turn enabled cold, dry, steppic conditions to persist in northern Siberia 20. However, this has been disproven as we know there was actually reduced sea ice cover during the last interglacial as a result of the warmer climate 21.
More recently, a new possible explanation for the extinction of mammoth steppe fauna may be emerging. Kienast et al. (2008) analyzed Eemian-age pollen deposits from the New Siberian Islands and found that dry herbaceous taxa were much more abundant than in the modern vegetation of the region 22. They argued that this was due to the Laptev Shelf being exposed to a much greater extent during the Eemian than during the Holocene, such that the New Siberian Islands remained connected to the Eurasian mainland despite globally higher sea levels. According to this hypothesis, the expanded shelf pushed the Arctic coastline much farther north and reduced maritime influence, resulting in a drier and more continental climate across northeastern Siberia during the last interglacial.
The Holocene encroachment of the Arctic shoreline-which occurred gradually over the course of the early to middle Holocene rather than immediately-was, according to the authors, the result of tectonic differences that caused the Laptev Shelf to sit lower than during the Eemian. The authors speculate that the marine transgression brought cooler, wetter summers to northeastern Siberia by increasing maritime influence, with the effects extending into other parts of northern Eurasia and possibly even Alaska.
The result was the dominance of typical moist and unpalatable tundra in the Holocene. In contrast, the more extensively exposed continental shelves of previous interglacials may have helped maintain drier conditions that allowed grassland communities to flourish. If correct, this could help explain why mammoths and other steppe fauna survived earlier interglacials but not the current one.
Could we finally have an answer to the question we’ve been asking for so long? It sounds convincing at first glance. In fact, scientists analyzing deposits from the Batagay thaw slump in inland Yakutia, not far from Verkhoyansk, found evidence that the Eemian interglacial supported somewhat more open vegetation than the Holocene in the region 23. One study linked this difference to the greater exposure of the continental shelf and the drier conditions it may have produced 24, suggesting-just as the Kienast et al. paper did-that the climatic influence of shelf exposure extended well beyond the immediate vicinity of the New Siberian Islands and Arctic coast.
But there are two glaring problems. The first is that the overall evidence for continental shelf exposure having sweeping effects on climate, via allegedly increasing aridity, on northeast Siberia (let alone beyond) is weak at the moment. The second is that the timeline and spatial pattern of mammoth extinction simply does not cleanly align with either this hypothesis, or other climate-linked hypotheses in general.
We will start with the first issue.
The Continental Shelf: How Influential Really?
A basic expectation of the notion that greater continental shelf exposure had wide-reaching as opposed to localized consequences would be that northeast Siberia and further afield would have been drier as a whole during previous interglacials. But also, recall that the early Holocene had a continental shelf exposure closer to the Eemian than to the late Holocene, with most of it still being dry land 25. Hence, we would also expect the early Holocene to be noticeably drier compared to the late Holocene.
We have already mentioned the pollen record from Lake Elgygytgyn in Chukotka, right next to Yakutia, which is extensively long and covers multiple past climate cycles. It indicates that interglacials of the past, such as the Eemian, were not only warmer, but also just as wet or wetter than the present one 17. Greater warmth, especially in summer, would be predicted by the continental shelf hypothesis of Kienast et al., but greater moisture would not be.
Pollen data from Lake Levinson in the Taymyr peninsula about 176 km from the Arctic Ocean revealed an early Holocene climate no drier than present26, despite the Laptev Shelf being much more strongly exposed then. The Yana site complex in Yakutia is located only about 80 km from the Arctic, and directly south of the submerged shelf. Yet, climatic reconstructions for the very early Holocene in Yana indicates that compared to present, summers were 0.8 C warmer but annual precipitation about 60 mm higher 27. This means that the slightly warmer summers were offset by increased precipitation, resulting in moisture levels similar to present. Moving inland, at Lake Temje near Yakutsk further south, chironomids suggest cooler temperatures than at present and a deeper lake level, the latter pointing to more humid conditions than present 28.
Hence, the data so far from sites in Taymyr, Yakutia, as well as Chukotka, suggests that whatever influence an extended continental shelf had on enhancing aridity was probably localized rather than widespread given that Eemian and/or early Holocene conditions were as or more humid than present. This casts doubt on the possibility that this shelf exposure had pervasive, important effects across northeast Siberia and beyond. The more open last interglacial vegetation inferred from the Batagay thaw slump remains curious, but we may speculate that it could have been the result of other factors such as megafaunal disturbance or greater wildfire activity due to warmer summers with more lightning rather than substantially higher aridity.
To a degree, this makes sense. The summer cooling influence of the Arctic Ocean drops off sharply moving 100 to 200 km inland and then levels off moving even further south 29. Hence, reduced evaporation over land due to Arctic marine transgression was unlikely to seriously influence areas that were already a safe distance from the coast to begin with. Further, the warm summers of the Eemian and early Holocene resulted in reduced Arctic sea ice 21 30, which may have resulted in more evaporation from the sea and more moisture reaching land and possibly offsetting the enhanced distance from sea compared to present.
But then there is the other issue, which is tying the timing of marine transgression, and really all proposed environmental changes, with the decline of woolly mammoths across their range.
Extinction: Still Too Much Steppe, Too Soon
Mammoths on the Eurasian mainland drop out of the fossil record entirely by the early Holocene 31. The most recent fossil of mainland mammoth is about 10,800 years old from what is now Zokhov Island 32, part of the New Siberian Island group that was still connected to Siberia by land at the time. This alone calls into question the speculation put forth by Kienast et al. about the importance of the Laptev Shelf in providing a haven for steppe-tundra animals, as the shelf would not become fully inundated until thousands of years after the last (currently known) mammoth lived there 25. Still, it is undeniable that due to a variety of climatic forcings, shrubs and trees did encroach on the mammoth steppe as the megafauna disappeared from the mainland, and we must address this.
But first, we need to ask a crucial question: at what level of tree and shrub cover do woolly mammoths, woolly rhinos, steppe bison, horses, etc. actually vanish from a given region? 80%? 90%? At what moisture index does their survival become untenable? No one really knows. It’s simply been assumed that all modern taigas and tundras are too woody and/or wet to support these creatures, because these environments are believed to not contain enough forage and also because during the glacial period, the animals overwhelmingly lived on open plains.

But there is some evidence that at least some megafauna did live in well-wooded environments, which is critical in ascertaining what caused them to disappear during the Pleistocene-Holocene transition and if very open steppes were really essential to their survival. The following discussion about habitat preferences and paleoenvironmental changes is admittedly rather dense, but it is an important read nonetheless.
Eastern mid-latitude North America is often ignored in discussions of woolly mammoth habitat. This part of the world was dominated by moist boreal forests and woodlands. During full glacial times, the mammoths lived as far south as Kentucky and Virginia 33 34. During late glacial times, they had moved into the Midwest and Northeast 35 36. These landscapes were usually a bit more open than most modern boreal forests but it may well be that the megafauna themselves were responsible for that as there is no evidence for eastern North America being particularly dry compared to the present during glacial times, and it most certainly was not drier than, for example, the modern taiga of Yakutia.
In Primorsky Krai in the southeast of Russia, woolly mammoths, woolly rhinos, horses, and steppe bison lived between 30 and 40 thousand years BP in a humid coastal environment that contained substantial tree cover in addition to steppe 37. Low nitrogen isotopic values of most of the herbivores’ bones testify to the humid climate and may indicate a partial incorporation of browse in their diets. These were not trapped, starving terminal populations. These animals lived near the Manchurian grasslands in the heart of the last glacial period, so their choice to live in these woodlands indicates they were suitable.
The persistence of mammoths and their compatriots in these well-wooded regions is indispensable context when investigating their disappearance during the last glacial-interglacial transition, a time when similar habitats were becoming established across northern Eurasia. It is certainly true that trees and shrubs proliferated heavily during the late glacial and early Holocene. However, these changes were neither instantaneous nor uniform. Some regions still retained substantial proportions of steppic taxa during the Bølling–Allerød and especially the Younger Dryas, and to an extent even the early Holocene (Preboreal).
Yet, when we look at fossil mammoth occurrences, there is a sharp decline everywhere south of the Arctic circle during the Bølling–Allerød (BA) warming 31 38 39. And by the early Holocene, the animals are almost entirely restricted to north of the Arctic Circle with the exception of some woolly mammoths living in close vicinity of the decaying ice sheet in eastern Europe. By the mid-Holocene, they only survived on Wrangel and Saint Paul Islands. The significance of all of this will be explained after we take a look at patterns of vegetative change.
Starting in the east: vegetation in central and to a lesser extent southern Yakutia was still rather steppic during the BA 40. Grass and forb (Artemisia) communities were plentiful, combined with a substantial component of both tree and shrub birch. The Younger Dryas (YD) saw reduced tree/shrub cover and an increase in herbs. The region around Lake Satagay in the western Yakutian lowlands seems to have had substantial herb cover with open woodlands even going into the early Holocene, possibly due to the heavy fire activity of that period 41. Hence, there was plenty of forage to feed mammoths and other large animals.
Even in the West Siberian Plain, today covered by an impenetrable expanse of marshy boreal forest, the late glacial transition period featured a mixed woody-herb cover as indicated by the pollen spectra at Entarny, located at 61° N 79° E 29. Pollen of non-arboreal taxa at Entarny hovered between 60-40% during the BA, YD, and much of the Preboreal, but mammoth populations in the West Siberian plain, like elsewhere had collapsed during the BA. The most recent mammoth was from a site known as Volchia Griva in the southern part of the plain, where mammoths lived until around 12,500 years ago during the Younger Dryas 32.
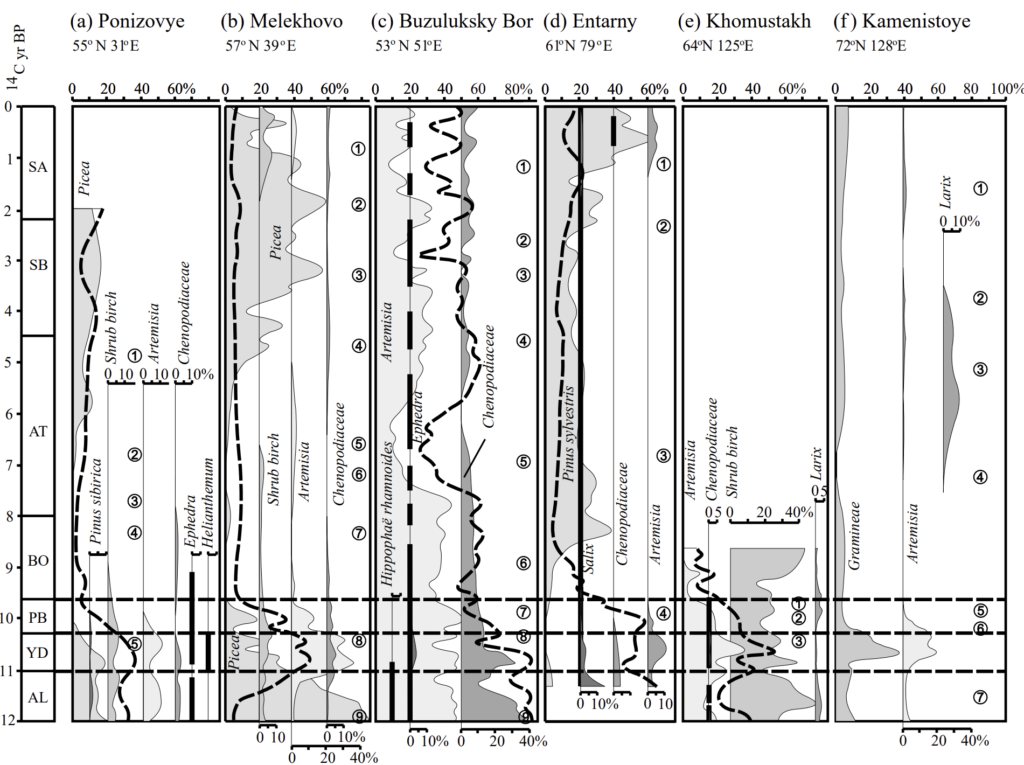
Moving to the East European Plain, a site known as Ponizovye in far western Russia documents another case in which dense forests were not fully established by the BA or YD with non-arboreal pollen falling within a healthy range of 30-40% 29. At Lake Chukhlomskoye a few hundred kilometers northeast of Moscow, the BA featured spruce-birch forests mixed in with periglacial steppe and meadow communities, and during the YD, periglacial birch forest-steppes dominated 42. Only during the early Holocene were the forest-steppes gradually replaced by proper forest.
Based on fossil occurrences, the last mainland Eurasian mammoths mostly survived north of the Arctic Circle in places such as the Yamal Peninsula, Taymyr, and the New Siberian Islands 32. These were regions dominated by tundra or steppe-tundra, where shrubs were of the dwarf variety and trees were scarce or absent altogether. Further south, the few mammoths that persisted into the early Holocene in places like Estonia and Cherepovets in Russia were living in or (with regard to Cherepovets) within migratory distance of a relict belt of periglacial steppe-tundra that hugged the decaying Fennoscandian ice sheet 43.
What is striking is that their final refugia were not in dry inland regions such as the Yana-Oymyakon highlands of Yakutia or the Ukok Plateau in the Altai-Sayan region, which might be expected under a primarily climate-driven extinction scenario. The latter region is often regarded as the closest modern analog to the mammoth steppe 44, making the absence of late-surviving mammoths there particularly puzzling.
Moreover, mammoth populations seem to have collapsed at the first sight of open boreal woodlands and forest-steppes spreading across Eurasia, even though temperatures remained fairly cool and grasses and forbs were still abundant in many areas. The shift toward Holocene-type environments appears to have been more gradual and regionally variable than is often implied, yet mammoths retreated at breakneck speed towards the few areas where trees remained sparse or absent, namely the Arctic and the remaining periglacial steppe-tundra zone in Eastern Europe.
One could be forgiven for looking at this trend and imagining woolly mammoths to be “tree-phobic.” But that view is difficult to reconcile with the fact that mammoths in eastern North America inhabited humid parklands, woodlands, and perhaps even forests 34 35 36, while mammoths, woolly rhinos, and steppe bison also persisted in the fairly wooded environments of Primorye between 30 and 40 thousand years BP 37. Taken together, the evidence suggests that cause of the disappearance of these animals was more complicated than a straightforward climate-driven environmental decline.
So what on Earth was actually happening?
No Escape
Wood is a critical resource for human beings. It was used to construct hunting weapons and settlements. Hunter-gatherers also used wood to make fire, without which they could not have survived the extremely harsh, lengthy winters of these latitudes. Fire provided warmth, enabled cooking, acted as a source of light during the long dark winter nights, and served as a defense against predators. There were alternative materials to make weapons, homes, and fire but obviously nothing was as practical as a readily available supply of wood, which of course would come with abundant trees or large shrubs.
During the colder and drier phases of the Late Pleistocene, human populations were much denser in the more climatically moderate southern portions of the mammoth steppe than in the far north 45 46, likely because woody vegetation was sparser there 47. The few people living in northern Siberia were probably concentrated near rivers or other localized areas where trees and large shrubs could still grow. Some researchers have even argued that the extreme conditions of the Last Glacial Maximum temporarily depopulated Siberia entirely 47 48, though this remains debated with others claiming continuous but reduced presence 46.
In this sense, the limiting factor for humans was not the absence of prey animals but rather the lack of wood as a consequence of the harsh climate 8. The very low density of humans across northern Siberia and areas bordering the ice sheets likely allowed the coldest parts of the mammoth steppe to function as a refugium for megafauna throughout much of the glacial period, allowing them to persist despite pervasive hunting in the more southerly areas where they overlapped substantially with humans.
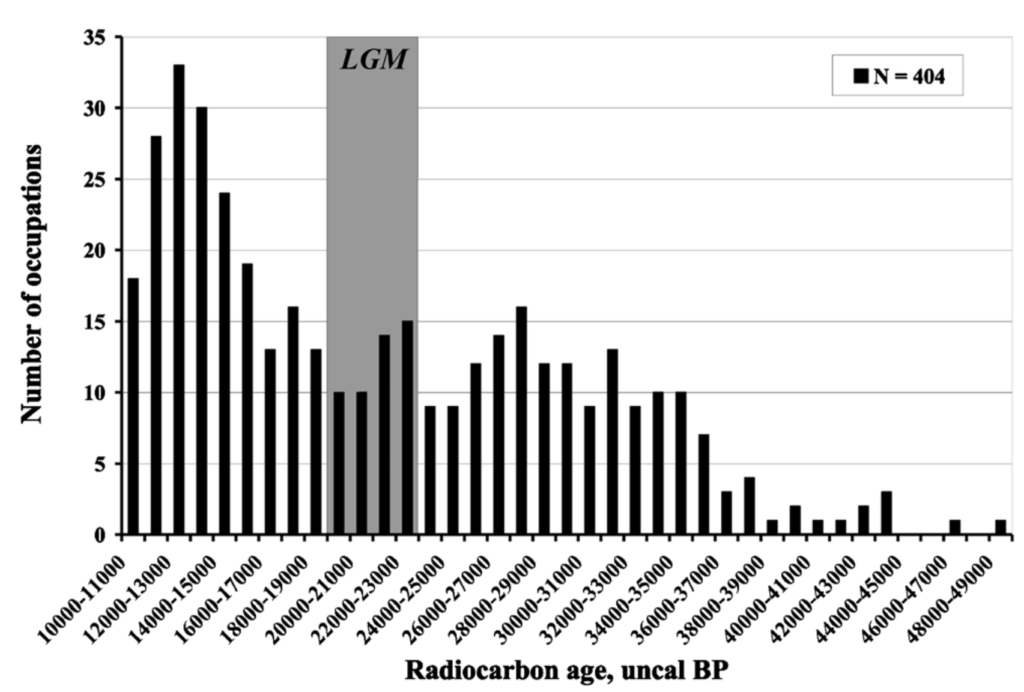
This all changed dramatically with late glacial warming and increased moisture, however. As large shrubs and trees surged in abundance in formerly very open steppe-tundra regions, humans were able to expand into areas that had previously supported only sparse occupation. During the Bølling–Allerød, forests still had not fully been established across much of northern Eurasia, but there was enough woody cover for humans to exploit. The archaeological evidence supports this notion, with the number of Siberian sites increasing sharply following the LGM 46.
Towards the end of the LGM, humans in Eurasia had also developed and spread microblade technology, which was highly effective for killing large animals and especially convenient for the highly mobile lifestyle of late glacial hunters 47 48. Dogs had also been domesticated by a number of cultures 49 50. Expanding human populations, armed with improved hunting technology and canines for assistance, may therefore have become a far more serious threat to mammoths and other megafauna than ever before.
Additionally, with the spread of woodland and shrubland during the late glacial, animals would have been forced to spend time in clearings in order to graze. This is in contrast to being widely dispersed across open steppe-tundra during the full glacial, as they had been previously. This circumstance would probably have made them particularly vulnerable to human hunters, who could reliably use such clearings to locate and target the animals. Additionally, the greater heterogeneity of the landscape and tree/shrub cover could have made it easier for humans to ambush animals 8.
Humans were already well-established across much of Eurasia, having adapted to tens of thousands of years of increasingly intense glacial conditions. Following the Last Glacial Maximum, even modest climatic amelioration was likely sufficient to enable their northward expansion. The cold, dry Younger Dryas may have temporarily slowed or reversed this conquest of the north, potentially allowing woolly mammoths to recover somewhat and then persist into the early Holocene in a few refugial areas. The next round of warming heading into the Holocene Thermal Maximum allowed trees and large shrubs to reach their northernmost limits51-even beyond where they are at present-which may have been the final nail in the coffin by allowing humans to expand further north than ever.
This could help explain why the last mainland Eurasian mammoths survived mainly in or near regions where trees remained scarce, such as the deep Arctic or areas adjacent to the Fennoscandian ice sheet. These regions were not only environmentally suitable for cold-adapted fauna, but also comparatively inaccessible to humans. By contrast, cool and dry inland areas such as the Yana-Oymyakon highlands or the Altai-Sayan, despite retaining substantial grazing habitat, likely supported higher human densities because wood was more readily available, and hence lost their mammoths earlier on.
In mid-continental North America, by contrast, human arrival occurred much later than in Eurasia, and their populations required time to grow and expand before they became a serious threat to the local animals. This delay might explain why woolly mammoths were flourishing in full glacial and late glacial woodland and forest environments there, but not in Eurasia where humans were an already entrenched and lethal force that rapidly sprang northwards thanks to deglacial warming and killed off the animals before they could settle into the newly-forming landscapes.
Mammoths have been the main star of today’s discussion, but a similar and perhaps even more striking pattern took place with woolly rhinos. The last recorded woolly rhinoceros from Europe dates not to the Bølling–Allerød, but earlier at around 15-16 thousand years ago during the Oldest Dryas 52. This was a time when conditions were still sufficiently cold, dry, and steppic on that continent 53 54, so environmental pathways for their extinction there are hard to muster. A better explanation could be the human Magdalenian and Epigravettian cultures, which had expanded northwards from their LGM refugia around this time 55 56. These highly adept and technologically advanced hunters may have been the end of the rhinos in Europe.
In northeast Siberia, woolly rhinos most likely disappeared during the Bølling–Allerød 52, yet one recent study by Guðjónsdóttir et al. (2025) found no evidence of inbreeding in early BA populations 57. Although Guðjónsdóttir et al. (2025) ignored a human role, the abruptness of the extinction during the BA contrasted with the still-healthy representation of herbaceous vegetation in the broader region in the same period 27 40 is less consistent with a climatic cause of extinction than an anthropogenic one. Rhinos could have been especially vulnerable to hunting due to their smaller populations, more limited mobility (due to shorter legs), and perhaps lower intelligence (due to smaller brains) compared to mammoths.

Meanwhile, in Alaska and the Yukon, it has been correctly pointed out in a study by Monteath et al. (2021) that shrub expansion-involving willows and birches-preceded extinctions of mammoth and horse 58. However, the first detectable evidence for human occupation in the region also appears shortly following the increase in shrubs. Woodland proper had not yet been established in the region 59, but warm summer temperatures during the late glacial 60 could have allowed the shrubs to grow quite tall and serve a similar functional role as trees for humans. Hence, declines in large herbivore carrying capacity due to shrub expansion alone are plausible, but drawing a direct line between this environmental shift and their extinctions is not advisable.
Reconsidering Ecological Adaptiveness
Ultimately, the fatal issue with climate-driven explanations is that there is no true control comparison. The transformation into woody Holocene-type environments across northern Eurasia occurred at the same time humans effectively colonized the far north with increasingly sophisticated hunting tools and dogs. The steppe-tundra fauna did not have a proper chance to “prove” their ability to thrive in the emerging landscapes.
This issue is compounded by the fact that we have few securely dated remains of these animals from earlier interglacials to give us a clear picture of how they responded to past Holocene-like environments when anatomically modern humans were absent. However, an extremely intriguing recent study looked into plant remains from the gastrointestinal tract of a woolly mammoth calf retrieved in 2024 from the Batagay thaw slump 61. The age of the calf, based on the surrounding sediments, suggests it dates to MIS 5e, also known as the Eemian interglacial.
The study revealed that the young animal lived in a taiga with light larch tree cover and substantial layer of birch, alder, and willow shrubs. It had a diverse diet with sedges (especially cotton-grass) being the most important with also smaller amounts of grass, forbs, and perhaps tree and shrub leaves. The mainly moisture-loving vegetation in its diet suggests it may have last grazed in a wetland area.
Although cotton-grass is said to have lower nutritional value than dryland taxa 62, the mammoths were clearly making do with it, assuming this calf was not an outlier and its diet reflects broader trends among contemporary individuals in the region. Additionally, cotton-grass is one of the most common flowering plants in the Arctic and Subarctic regions today.
In fact, even during glacial times, mammoth individuals from northeast and northwest Siberia were found to be eating primarily grasses and sedges, which are still common in northerly areas today despite the comparatively large proportions of shrub and moss 63 64. And similar gastrointestinal analysis of the steppe bison has shown it was a mixed feeder whose diet included forbs, graminoids, and shrubs 65. All of this has optimistic implications for the potential ability of woolly mammoths and similar fauna to have survived well into the Holocene in the absence of human interference.
Even so, we can still say with confidence that warming alone would have reduced the geographic distribution of woolly mammoths (and woolly rhinos), as they were, after all, cold-adapted and the Arctic/Subarctic climate zones shrank in size. The reduction in the abundance of total herb forage due to humidification most likely would have also lowered megafaunal population densities in areas they still occupied. However, reduced densities would not necessarily have meant extinction. Even today, the Arctic and Subarctic remain vast expanses, and in the absence of sustained human pressure, substantial populations of these animals may well have persisted.
What is the crucial point to take away from this? It’s that the answer to the question of how these animals survived previous interglacials will not be answered via identifying cryptic “steppe-tundra” refugia that existed due to the peculiar climatic characteristics (compared to the Holocene) of said past interglacials. Rather, the dilemma will be solved by recognizing and appreciating the ability of the animals to persevere under ecological conditions similar to those that exist today.
Megafauna Impacts on Environments
What impacts could the megafauna have had on modern ecosystems if they had not gone extinct, and what can modern rewilding hope to accomplish? First, it is important to recall that the mammoth steppe, in its strict sense, was a largely treeless environment. Treeless environments occur when it is either too cold or too dry for tree growth, and realistically no level of herbivore pressure alone could convert modern boreal forests into true steppe-tundra, given the warm summers and sufficient moisture. That said, sustained grazing and disturbance could plausibly shift parts of the taiga toward more mosaic conditions. These might be similar to the Pleistocene boreal parklands that supported diverse fauna, including the extinct giant deer Megaloceros 66.
The Arctic tundra presents a different case, since low summer temperatures directly limit tree growth regardless of biotic influences. Here, large herbivores could potentially reduce shrub, moss, and lichen dominance while favoring grasses and forbs, producing landscapes partially analogous to steppe-tundra. However, Pleistocene rewilding has not been extensively conducted in tundras so far aside from a few reintroductions of reindeer and muskoxen, so more research is needed.
Importantly, the magnitude of herbivore-driven change depends strongly on local conditions. For instance, Reinecke et al. (2021) found that in Zimov’s Pleistocene Park, herbivore impacts were relatively modest, likely due to the site’s more maritime-influenced and humid climate 67. In contrast, a bison enclosure in central Yakutia showed far stronger shifts toward vegetative openness and increased soil aridity under herbivore pressure, reflecting its more continental, drier setting. This suggests that herbivores are most effective as ecosystem engineers in relatively dry environments.
The study also notes that there are no modern analogs of the woolly mammoth and woolly rhino at high latitudes. These were two megaherbivores far heavier than the largest animals at Pleistocene Park, the former having trunks and tusks and the latter with a large horn, builds that are ideal for impacting vegetation. As such, modern-day rewilding efforts such as at Pleistocene Park may be less effective at altering landscapes compared to a scenario where the extinct members of the mammoth steppe fauna had still existed.

It is difficult to know exactly what modern boreal and Arctic ecosystems would look like if the fauna had not disappeared, but the REVEALS method of pollen reconstruction could offer a useful framework for testing such possibilities, as it corrects for biases in pollen representation and has been used to analyze past ecosystems at high resolutions 68. This approach has already shown that last interglacial landscapes in parts of Europe were more heterogeneous and open than previously assumed 69.
REVEALS remains underutilized in Siberia, Alaska, and the Yukon, but future work using this method on past interglacial pollen deposits, along with extensively sampling more sites, may indicate a similar pattern after controlling for the effects of climate and fire. We could discover that during interglacials, cold-adapted megafauna had similar effects on the aforementioned regions as temperate European fauna did in Europe. And we could speculate that if they never went extinct, modern ecosystems such as certain boreal forests may be more open woodland or parkland-like, while some tundras might resemble a sort of quasi steppe-tundra.
A separate question is whether such ecological changes would meaningfully affect carbon storage, permafrost stability, or climate, as suggested by Zimov 8 9. Even under optimistic scenarios, it is difficult to envision a return to huge swathes of steppe-tundra under modern climatic conditions. The most likely outcome would be an increase in habitat mosaics and modestly greater openness rather than wholesale biome replacement, meaning any albedo-driven cooling effects would be limited and we would not see substantially increased permafrost resilience.
This is consistent with what we know of past interglacials, which were as warm or warmer than the Holocene and supported roughly similar levels of tree and shrub cover 15 17. While large herbivores may have influenced vegetation structure locally, there is little evidence that they fundamentally altered large-scale carbon storage or climate trajectories and the same would be true for herbivores utilized for modern rewilding projects.
Conclusion
Ultimately, the awe-inspiring mammoth steppe, despite how well-studied it is, is a complex and confusing topic. The biggest controversy and lack of clarity surrounds the cause of the extinction of many of its iconic animals, such as woolly mammoths, woolly rhinos, steppe bison, cave lions, and more. Despite a substantial number of scientific papers (including many meticulous and insightful ones) arguing that climate-driven environmental changes were solely or mainly responsible for their loss, the reality is probably more sobering and involves a major and necessary anthropogenic component as well.
To summarize what my article here argues:
- The mammoth steppe in the strict sense, i.e. a treeless biome, was formed and maintained mainly by climate and other abiotic factors. Herbivores may have enriched it through their activities but they did not create it.
- It largely disappeared due to climatic shifts, consistent with patterns seen in earlier interglacials.
- This climatic transition did not necessarily imply inevitable extinction of the fauna, which were ecologically flexible and could most likely make do with and continue to enrich the newly forming habitats.
- Population decline and extinction occurred too rapidly to be ascribable wholly or predominantly to environmental change, as transitional habitats were still relatively open and contained plenty of forage for megafauna.
- Human expansion into northern Eurasia after the LGM was facilitated by the spread of woody vegetation, which increased resource availability and allowed wider settlement. The geographical extent of human settlement and lethality of human hunters was unprecedented compared to earlier in the glacial, which is a confounding factor when attempting to disentangle the roles of humans versus climate in extinction.
- Treeless refugial habitats with low or no human presence contracted over time with warming, eventually dooming woolly mammoths and company.
- In the absence of human pressure, now-extinct species likely would have persisted, albeit at reduced ranges and densities.
- Megafaunal ecosystem engineering may have produced important but regional and site-specific, not continental-scale, environmental changes.
- This means that current rewilding, even if able to be done at mass scale, is unlikely to be effective at mitigating permafrost melt and climate change.
Overall, the goal here is to emphasize that ecological change, and animal responses to it, are more complex than many appear to believe, and that correlation is insufficient to assume causation, particularly when confounding variables are involved. The viewpoints and facts presented here could provide important context and framework for future research.
References
- Guthrie, R. D. (1990). Frozen fauna of the Mammoth Steppe : the story of the blue babe. University Of Chicago Press.[↩]
- Yurtsev, B. A. (2001). The Pleistocene “Tundra-Steppe” and the productivity paradox: the landscape approach. Quaternary Science Reviews, 20(1-3), 165–174. https://doi.org/10.1016/s0277-3791(00)00125-6[↩][↩]
- Kahlke, R.-D. (2014). The origin of Eurasian Mammoth Faunas (Mammuthus–Coelodonta Faunal Complex). Quaternary Science Reviews, 96, 32–49. https://doi.org/10.1016/j.quascirev.2013.01.012[↩][↩]
- Dale Guthrie, R. (2001). Origin and causes of the mammoth steppe: a story of cloud cover, woolly mammal tooth pits, buckles, and inside-out Beringia. Quaternary Science Reviews, 20(1-3), 549–574. https://doi.org/10.1016/s0277-3791(00)00099-8[↩][↩][↩][↩]
- Zimov, S. A., Zimov, N. S., Tikhonov, A. N., & Chapin, F. S. (2012). Mammoth steppe: a high-productivity phenomenon. Quaternary Science Reviews, 57, 26–45. https://doi.org/10.1016/j.quascirev.2012.10.005[↩]
- Willerslev, E., Davison, J., Moora, M., Zobel, M., Coissac, E., Edwards, M. E., Lorenzen, E. D., Vestergård, M., Gussarova, G., Haile, J., Craine, J., Gielly, L., Boessenkool, S., Epp, L. S., Pearman, P. B., Cheddadi, R., Murray, D., Bråthen, K. A., Yoccoz, N., & Binney, H. (2014). Fifty thousand years of Arctic vegetation and megafaunal diet. Nature, 506(7486), 47–51. https://doi.org/10.1038/nature12921.[↩]
- Zimov, S. A., Chuprynin, V. I., Oreshko, A. P., Chapin, F. S., Reynolds, J. F., & Chapin, M. C. (1995). Steppe-Tundra Transition: A Herbivore-Driven Biome Shift at the End of the Pleistocene. The American Naturalist, 146(5), 765–794. https://doi.org/10.1086/285824[↩]
- Zimov, S. A., Zimov, N. S., Chapin, F. S., Iii, & Zimov, S. A. (2012). The Past and Future of the Mammoth Steppe Ecosystem. Paleontology in Ecology and Conservation. https://doi.org/10.1007/978-3-642-25038-5_10[↩][↩][↩][↩]
- Zimov, S. A. (2005). Pleistocene Park: Return of the Mammoth’s Ecosystem. Science, 308(5723), 796–798. https://doi.org/10.1126/science.1113442[↩][↩]
- Stewart, M., Peters, C., Ziegler, M. J., Carleton, W. C., Roberts, P., Boivin, N., & Groucutt, H. S. (2025). The state of the late Quaternary megafauna extinction debate: a systematic review and analysis. Frontiers in Mammal Science, 4. https://doi.org/10.3389/fmamm.2025.1678231[↩]
- Gerhart, L. M., & Ward, J. K. (2010). Plant responses to low CO2 of the past. New Phytologist, 188(3), 674–695. https://doi.org/10.1111/j.1469-8137.2010.03441.x[↩]
- Walker, D. A., Bockheim, J. G., Chapin III, F. S., Eugster, W., Nelson, F. E., & Ping, C. L. (2001). Calcium-rich tundra, wildlife, and the “Mammoth Steppe.” Quaternary Science Reviews, 20(1-3), 149–163. https://doi.org/10.1016/s0277-3791(00)00126-8[↩]
- Kaplan, J. O., Pfeiffer, M., Kolen, J. C. A., & Davis, B. A. S. (2016). Large Scale Anthropogenic Reduction of Forest Cover in Last Glacial Maximum Europe. PLOS ONE, 11(11), e0166726. https://doi.org/10.1371/journal.pone.0166726[↩]
- Mahowald, N., Kohfeld, K., Hansson, M., Balkanski, Y., Harrison, S. P., Prentice, I. C., Schulz, M., & Rodhe, H. (1999). Dust sources and deposition during the last glacial maximum and current climate: A comparison of model results with paleodata from ice cores and marine sediments. Journal of Geophysical Research: Atmospheres, 104(D13), 15895–15916. https://doi.org/10.1029/1999jd900084[↩]
- Tarasov, P., Granoszewski, W., Bezrukova, E., Brewer, S., Nita, M., Abzaeva, A., & Oberhänsli, H. (2005). Quantitative reconstruction of the last interglacial vegetation and climate based on the pollen record from Lake Baikal, Russia. Climate Dynamics, 25(6), 625–637. https://doi.org/10.1007/s00382-005-0045-0[↩][↩]
- Prokopenko, A. A., Bezrukova, E. V., Khursevich, G. K., Solotchina, E. P., Kuzmin, M. I., & Tarasov, P. E. (2010). Climate in continental interior Asia during the longest interglacial of the past 500 000 years: the new MIS 11 records from Lake Baikal, SE Siberia. Climate of the Past, 6(1), 31–48. https://doi.org/10.5194/cp-6-31-2010[↩]
- Tarasov, P. E., Andreev, A. A., Anderson, P. M., Lozhkin, A. V., Leipe, C., Haltia, E., Nowaczyk, N. R., Wennrich, V., Brigham-Grette, J., & Melles, M. (2013). A pollen-based biome reconstruction over the last 3.562 million years in the Far East Russian Arctic – new insights into climate–vegetation relationships at the regional scale. Climate of the Past, 9(6), 2759–2775. https://doi.org/10.5194/cp-9-2759-2013[↩][↩][↩]
- Muhs, D. R., Ager, T. A., & Begét, J. E. (2001). Vegetation and paleoclimate of the last interglacial period, central Alaska. Quaternary Science Reviews, 20(1-3), 41–61. https://doi.org/10.1016/s0277-3791(00)00132-3[↩]
- Schweger, C., Froese, D., White, J. M., & Westgate, J. A. (2011). Pre-glacial and interglacial pollen records over the last 3 Ma from northwest Canada: Why do Holocene forests differ from those of previous interglaciations? Quaternary Science Reviews, 30(17-18), 2124–2133. https://doi.org/10.1016/j.quascirev.2011.01.020[↩]
- Putshkov, P.V., (2003) – The impact of mammoths on their biome: clash of two paradigms – in:Reumer, J.W.F., de Vos, J. & Mol, D. (eds.) – ADVANCES IN MAMMOTH RESEARCH (Proceedings of the Second International Mammoth Conference, Rotterdam, Published 24 May 2003[↩]
- CAPE-Last Interglacial Project Members. (2006). Last Interglacial Arctic warmth confirms polar amplification of climate change. Quaternary Science Reviews, 25(13-14), 1383–1400. https://doi.org/10.1016/j.quascirev.2006.01.033[↩][↩]
- Kienast, F., Tarasov, P., Schirrmeister, L., Grosse, G., & Andreev, A. A. (2008). Continental climate in the East Siberian Arctic during the last interglacial: Implications from palaeobotanical records. Global and Planetary Change, 60(3-4), 535–562. https://doi.org/10.1016/j.gloplacha.2007.07.004[↩]
- Ashastina, K., Schirrmeister, L., Fuchs, M., & Kienast, F. (2017). Palaeoclimate characteristics in interior Siberia of MIS 6–2: first insights from the Batagay permafrost mega-thaw slump in the Yana Highlands. Climate of the Past, 13(7), 795–818. https://doi.org/10.5194/cp-13-795-2017[↩]
- Jongejans, L. L., Mangelsdorf, K., Karger, C., Opel, T., Wetterich, S., Courtin, J., Meyer, H., Kizyakov, A. I., Grosse, G., Shepelev, A. G., Syromyatnikov, I. I., Fedorov, A. N., & Strauss, J. (2022). Molecular biomarkers in Batagay megaslump permafrost deposits reveal clear differences in organic matter preservation between glacial and interglacial periods. The Cryosphere, 16(9), 3601–3617. https://doi.org/10.5194/tc-16-3601-2022[↩]
- Bauch, H. A., Mueller-Lupp, T., Taldenkova, E., Spielhagen, R. F., Kassens, H., Grootes, P. M., Thiede, J., Heinemeier, J., & Petryashov, V. V. (2001). Chronology of the Holocene transgression at the North Siberian margin. Global and Planetary Change, 31(1-4), 125–139. https://doi.org/10.1016/s0921-8181(01)00116-3[↩][↩]
- Lenz, M., Lenz, M. M., Andreev, A., Scheidt, S., Gromig, R., Lebas, E., Fedorov, G., Krastel, S., Melles, M., & Wagner, B. (2021). Climate and environmental history at Lake Levinson‐Lessing, Taymyr Peninsula, during the last 62 kyr. Journal of Quaternary Science, 37(5), 836–850. https://doi.org/10.1002/jqs.3384[↩]
- Pavlova, E. Y., Dorozhkina, M. V., Basilyan, A. E., & Pitulko, V. V. (2026). High resolution record of vegetation and climate change in Western Beringia (Yana–Indighirka Lowland) at the end of the Late Pleistocene. Arctic Antarctic and Alpine Research, 58(1). https://doi.org/10.1080/15230430.2026.2627696[↩][↩]
- Nazarova, L., Lüpfert, H., Subetto, D., Pestryakova, L., & Diekmann, B. (2013). Holocene climate conditions in central Yakutia (Eastern Siberia) inferred from sediment composition and fossil chironomids of Lake Temje. Quaternary International, 290-291, 264–274. https://doi.org/10.1016/j.quaint.2012.11.006[↩]
- Velichko, A. A., Catto, N., Drenova, A. N., Klimanov, V. A., Kremenetski, K. V., & Nechaev, V. P. (2002). Climate changes in East Europe and Siberia at the Late glacial–holocene transition. Quaternary International, 91(1), 75–99. https://doi.org/10.1016/s1040-6182(01)00104-5[↩][↩][↩]
- Dyck, S., Tremblay, L. B., & de Vernal, A. (2010). Arctic sea-ice cover from the early Holocene: the role of atmospheric circulation patterns. Quaternary Science Reviews, 29(25-26), 3457–3467. https://doi.org/10.1016/j.quascirev.2010.05.008[↩]
- Kuzmin, Y. V. (2010). Extinction of the woolly mammoth (Mammuthus primigenius) and woolly rhinoceros (Coelodonta antiquitatis) in Eurasia: Review of chronological and environmental issues. Boreas, 39(2), 247–261. https://doi.org/10.1111/j.1502-3885.2009.00122.x[↩][↩]
- Kuzmin, Y., Dobretsov, N. N., & Lyamina, V. A. (2026). Spatiotemporal patterns of existence and extinction for woolly mammoth in Siberia in the last 50,000 years. Radiocarbon, 1–9. https://doi.org/10.1017/rdc.2026.10197[↩][↩][↩]
- Enk, J., Devault, A., Widga, C., Saunders, J., Szpak, P., Southon, J., Rouillard, J.-M., Shapiro, B., Golding, G. B., Zazula, G., Froese, D., Fisher, D. C., MacPhee, R. D. E., & Poinar, H. (2016). Mammuthus Population Dynamics in Late Pleistocene North America: Divergence, Phylogeography, and Introgression. Frontiers in Ecology and Evolution, 4. https://doi.org/10.3389/fevo.2016.00042[↩]
- Schubert, B. W., & Wallace, S. C. (2009). Late Pleistocene giant short-faced bears, mammoths, and large carcass scavenging in the Saltville Valley of Virginia, USA. Boreas, 38(3), 482–492. https://doi.org/10.1111/j.1502-3885.2009.00090.x[↩][↩]
- Saunders, J. J., Grimm, E. C., Widga, C. C., Campbell, G. D., Curry, B. B., Grimley, D. A., Hanson, P. R., McCullum, J. P., Oliver, J. S., & Treworgy, J. D. (2010). Paradigms and proboscideans in the southern Great Lakes region, USA. Quaternary International, 217(1-2), 175–187. https://doi.org/10.1016/j.quaint.2009.07.031[↩][↩]
- Feranec, R. S., & Kozlowski, A. L. (2016). Implications of a Bayesian radiocarbon calibration of colonization ages for mammalian megafauna in glaciated New York State after the Last Glacial Maximum. Quaternary Research, 85(2), 262–270. https://doi.org/10.1016/j.yqres.2016.01.003[↩][↩]
- Ma, J., Wang, Y., Baryshnikov, G. F., Drucker, D. G., McGrath, K., Zhang, H., Bocherens, H., & Hu, Y. (2021). The Mammuthus-Coelodonta Faunal Complex at its southeastern limit: A biogeochemical paleoecology investigation in Northeast Asia. Quaternary International, 591, 93–106. https://doi.org/10.1016/j.quaint.2020.12.024[↩][↩]
- Stuart, A. J. (2005). The extinction of woolly mammoth (Mammuthus primigenius) and straight-tusked elephant (Palaeoloxodon antiquus) in Europe. Quaternary International, 126-128, 171–177. https://doi.org/10.1016/j.quaint.2004.04.021[↩]
- Markova, A. K., Puzachenko, A. Yu., van Kolfschoten, T., van der Plicht, J., & Ponomarev, D. V. (2013). New data on changes in the European distribution of the mammoth and the woolly rhinoceros during the second half of the Late Pleistocene and the early Holocene. Quaternary International, 292, 4–14. https://doi.org/10.1016/j.quaint.2012.11.033[↩]
- Andreev, A. A., Klimanov, V. A., & Sulerzhitsky, L. D. (1997). Younger Dryas pollen records from central and southern Yakutia. Quaternary International, 41-42, 111–117. https://doi.org/10.1016/s1040-6182(96)00042-0[↩][↩]
- Ramesh Glückler, Geng, R., Grimm, L., Izabella Baisheva, Herzschuh, U., Stoof‐Leichsenring, K. R., Kruse, S., Andreev, A., Pestryakova, L. A., & Dietze, E. (2022). Holocene wildfire and vegetation dynamics in Central Yakutia, Siberia, reconstructed from lake-sediment proxies. Frontiers in Ecology and Evolution, 10. https://doi.org/10.3389/fevo.2022.962906[↩]
- Samus, A. V., Konstantinov, E. A., & Filippova, K. G. (2024). Late Glacial vegetation changes in the Kostroma Volga region based on pollen study of Lake Chukhlomskoye bottom sediments. Limnology and Freshwater Biology, 4, 642–647. https://doi.org/10.31951/2658-3518-2024-a-4-642[↩]
- Krivokorin, I., Poska, A., Vassiljev, J., Veski, S., & Amon, L. (2025). Environment of European Last Mammoths: Reconstructing the Landcover of the Eastern Baltic Area at the Pleistocene/Holocene Transition. Land, 14(1), 178. https://doi.org/10.3390/land14010178[↩]
- Chytrý, M., Horsák, M., Jiří Danihelka, Nikolai Ermakov, German, D. A., Hájek, M., Hájková, P., Kočí, M., Svatava Kubešová, Pavel Lustyk, Nekola, J. C., Věra Pavelková Řičánková, Zdenka Preislerová, Philipp Resl, & Valachovič, M. (2018). A modern analogue of the Pleistocene steppe-tundra ecosystem in southern Siberia. Boreas, 48(1), 36–56. https://doi.org/10.1111/bor.12338[↩]
- Tallavaara, M., Luoto, M., Korhonen, N., Järvinen, H., & Seppä, H. (2015). Human population dynamics in Europe over the Last Glacial Maximum. Proceedings of the National Academy of Sciences, 112(27), 8232–8237. https://doi.org/10.1073/pnas.1503784112[↩]
- Kuzmin, Y. V., & Keates, S. G. (2016). Siberia and neighboring regions in the Last Glacial Maximum: did people occupy northern Eurasia at that time? Archaeological and Anthropological Sciences, 10(1), 111–124. https://doi.org/10.1007/s12520-016-0342-z[↩][↩][↩]
- Goebel, T. (1999). Pleistocene human colonization of Siberia and peopling of the Americas: An ecological approach. Evolutionary Anthropology: Issues, News, and Reviews, 8(6), 208–227. https://doi.org/10.1002/(sici)1520-6505(1999)8:6<208::aid-evan2>3.0.co;2-m[↩][↩][↩]
- Goebel, T. (2002). The “microblade adaptation” and recolonization of Siberia during the Late Upper Pleistocene. Archeological Papers of the American Anthropological Association, 12(1), 117–131.[↩][↩]
- Marsh, W. A., Scarsbrook, L., Yüncü, E., Hodgson, L., Lin, A. T., De Iorio, M., Thalmann, O., Thomas, M. G., Goor, M., Bergström, A., Noseda, A., Amiri, S., Biglari, F., Borić, D., Bougiouri, K., Carmagnini, A., Giannì, M., Higham, T., Lebrasseur, O., & Linderholm, A. (2026). Dogs were widely distributed across western Eurasia during the Palaeolithic. Nature, 651(8107), 995–1003. https://doi.org/10.1038/s41586-026-10170-x[↩]
- Perri, A. R., Feuerborn, T. R., Frantz, L. A. F., Larson, G., Malhi, R. S., Meltzer, D. J., & Witt, K. E. (2021). Dog Domestication and the Dual Dispersal of People and Dogs into the Americas. Proceedings of the National Academy of Sciences, 118(6). https://doi.org/10.1073/pnas.2010083118[↩]
- Makeyev, V. M., Ponomareva, D. P., Pitulko, V. V., Chernova, G. M., & Solovyeva, D. V. (2003). Vegetation and Climate of the New Siberian Islands for the Past 15,000 Years. Arctic, Antarctic, and Alpine Research, 35(1), 56–66.[↩]
- Stuart, A. J., & Lister, A. M. (2012). Extinction chronology of the woolly rhinoceros Coelodonta antiquitatis in the context of late Quaternary megafaunal extinctions in northern Eurasia. Quaternary Science Reviews, 51, 1–17. https://doi.org/10.1016/j.quascirev.2012.06.007[↩][↩]
- Clark, P. U., Shakun, J. D., Baker, P. A., Bartlein, P. J., Brewer, S., Brook, E., Carlson, A. E., Cheng, H., Kaufman, D. S., Liu, Z., Marchitto, T. M., Mix, A. C., Morrill, C., Otto-Bliesner, B. L., Pahnke, K., Russell, J. M., Whitlock, C., Adkins, J. F., Blois, J. L., & Clark, J. (2012). Global climate evolution during the last deglaciation. Proceedings of the National Academy of Sciences, 109(19), E1134–E1142. https://doi.org/10.1073/pnas.1116619109[↩]
- Duprat-Oualid, F., Bégeot, C., Peyron, O., Rius, D., Millet, L., & Magny, M. (2022). High-frequency vegetation and climatic changes during the Lateglacial inferred from the Lapsou pollen record (Cantal, southern Massif Central, France). Quaternary International, 636, 69–80. https://doi.org/10.1016/j.quaint.2022.04.012[↩]
- Maier, A. (2015). The Recolonization of Central Europe. Vertebrate Paleobiology and Paleoanthropology, 231–241. https://doi.org/10.1007/978-94-017-7206-8_8[↩]
- Wiśniewski, A., Połtowicz-Bobak, M., Bobak, D., Jary, Z., & Moska, P. (2017). The Epigravettian and the Magdalenian in Poland: New chronological data and an old problem. Geochronometria, 44(1), 16–29. https://doi.org/10.1515/geochr-2015-0052[↩]
- Guðjónsdóttir, S. M., Lord, E., Pochon, Z., Lemež, Š., Dussex, N., Stanton, D. W. G., Sinding, M.-H. S., Fedorov, S., Dalén, L., & Chacón-Duque, J. C. (2025). Genome Shows no Recent Inbreeding in Near-Extinction Woolly Rhinoceros Sample Found in Ancient Wolf’s Stomach. Genome Biology and Evolution, 18(1). https://doi.org/10.1093/gbe/evaf239[↩]
- Monteath, A. J., Gaglioti, B. V., Edwards, M. E., & Froese, D. (2021). Late Pleistocene shrub expansion preceded megafauna turnover and extinctions in eastern Beringia. Proceedings of the National Academy of Sciences, 118(52). https://doi.org/10.1073/pnas.2107977118[↩]
- Dyke, A. S. (2007). Late Quaternary Vegetation History of Northern North America Based on Pollen, Macrofossil, and Faunal Remains*. Paleoenvironments, 59(2-3), 211–262. https://doi.org/10.7202/014755ar[↩]
- Kurek, J., Cwynar, L. C., Ager, T. A., Abbott, M. B., & Edwards, M. E. (2009). Late Quaternary paleoclimate of western Alaska inferred from fossil chironomids and its relation to vegetation histories. Quaternary Science Reviews, 28(9-10), 799–811. https://doi.org/10.1016/j.quascirev.2008.12.001[↩]
- Zanina, O. G., Lopatina, D. A., Novgorodov, G. P., & Cheprasov, M. Yu. (2026). Late Neopleistocene Vegetation and Environments of the Mammoth Calf Yana (Mammuthus primigenius (Blumenbach, 1799)) in the Batagay Section (Yakutia) Based on Microfossil Data. Doklady Earth Sciences, 527(2). https://doi.org/10.1134/s1028334x25610107[↩]
- Ukraintseva, V. V. (1981). Vegetation of warm late pleistocene intervals and the extinction of some large herbivorous mammals. Polar Geography and Geology, 5(4), 189–203. https://doi.org/10.1080/10889378109388689[↩]
- Kirillova, I. V., Argant, J., Lapteva, E. G., Korona, O. M., van, Zinovyev, E. V., Kotov, A. A., Chernova, O. F., Fadeeva, E. O., Baturina, O. A., Kabilov, M. R., Shidlovskiy, F. K., & Zanina, O. G. (2016). The diet and environment of mammoths in North-East Russia reconstructed from the contents of their feces. Quaternary International, 406, 147–161. https://doi.org/10.1016/j.quaint.2015.11.002[↩]
- Kosintsev, P. A., Lapteva, E. G., Korona, O. M., & Zanina, O. G. (2012). Living environments and diet of the Mongochen mammoth, Gydan Peninsula, Russia. Quaternary International, 276-277, 253–268. https://doi.org/10.1016/j.quaint.2011.11.004[↩]
- Jin, C., Forsythe, A., Miklós Bálint, Kienast, F., Nikolskiy, P. A., Kowalczyk, R., Katerina Guschanski, & Tom. (2025). Metagenomic insights into the diet and past Kolyma Lowland habitat from the intestinal content of a Late Pleistocene steppe bison. Quaternary Science Reviews, 369, 109604–109604. https://doi.org/10.1016/j.quascirev.2025.109604[↩]
- Lister, A. M., & Stuart, A. J. (2019). The extinction of the giant deer Megaloceros giganteus (Blumenbach): New radiocarbon evidence. Quaternary International, 500, 185–203. https://doi.org/10.1016/j.quaint.2019.03.025[↩]
- Reinecke, J., Ashastina, K., Kienast, F., Troeva, E., & Wesche, K. (2021). Effects of large herbivore grazing on relics of the presumed mammoth steppe in the extreme climate of NE-Siberia. Scientific Reports, 11(1), 12962. https://doi.org/10.1038/s41598-021-92079-1[↩]
- Sugita, S. (2007). Theory of quantitative reconstruction of vegetation I: pollen from large sites REVEALS regional vegetation composition. The Holocene, 17(2), 229–241. https://doi.org/10.1177/0959683607075837[↩]
- Pearce, E. A., Mazier, F., Normand, S., Fyfe, R., Andrieu, V., C.C. Bakels, Balwierz, Z., Krzysztof Bińka, Boreham, S., Borisova, O., Broström, A., Jacques‐Louis de Beaulieu, Gao, C., Penélope González-Sampériz, Wojciech Granoszewski, Hrynowiecka, A., Piotr Kołaczek, Petr Kuneš, Magri, D., & Malkiewicz, M. (2023). Substantial light woodland and open vegetation characterized the temperate forest biome before Homo sapiens. Science Advances, 9(45). https://doi.org/10.1126/sciadv.adi9135[↩]